

Аннотация
Introduction. Mercury and its compounds are among the most dangerous toxic substances, which makes mercury pollution one of the most urgent environmental issues. The present research objective was to study the accumulation of mercury and its impact on the terrestrial ecosystems in the area of the Beloosipovo mercury deposit (Kemerovo Region, West Siberia, Russia).Study objects and methods. The study used standard methods to test soil, herbs, herpetobiont insects, and small mammals. The sampling was conducted at 13 points in the cardinal directions at 0.5, 1.5, and 3.0 km from the pollution source. The method of atomic absorption was employed to measure the concentration of mercury in the samples prepared by the wet mineralization method.
Results and discussion. The main components of terrestrial ecosystems revealed no excessive concentration of mercury in the soil. However, the water samples from the Belaya Osipova river demonstrated an excess in the maximum permissible concentration of mercury from 5 to 20% (0.00056–0.00074 mg/L). Further up the food chains, the concentration of mercury in organisms decreased by 1–2 orders of magnitude, depending on the sampling point. The study also revealed Siberian trout lily (Erythronium sibiricum (Fisch. et C. A. Mey) Kryl.), which is protected at the federal and regional levels, as well as several nemoral tertiary relics.
Conclusion. The decreasing concentration of mercury in the food chains means the ecosystem is under no severe negative impact.
Ключевые слова
Ecology, mercury, mercury-containing compounds, terrestrial ecosystems, food chainsВВЕДЕНИЕ
Mercury is an important safety issue in the environmental, medical, and social aspects. In fact, mercury-related issues are one of the most urgent contemporary challenges. Mercury (Hg) and mercurycontaining compounds are toxic substances that pose danger to all living organisms. According to preliminary estimates, about 4700 tons of mercury is discharged worldwide every year [1–3]. Mercury-related water pollution is especially dangerous, since water-soluble toxic methylmercury [CH3Hg] accumulates in the fish as a result of activity of sediment microorganisms.
Mercury affects land and water plants, animals, fungi, and microorganisms, which constantly interact with each other in food chains, symbiosis, and etc. [4]. Many studies recognize the essential role of terrestrial plants in the biogeochemical cycle of mercury [5–7]. For instance, Leonard et al. tested five plant species for absorption, distribution, and subsequent release of mercury into the atmosphere, namely Lepidium latifolium L., Artemisia douglasiana Bessin Hook, Caulanthus sp. Watson, Fragaria vesca L., and Eucalyptus globulus Labill [8]. The research featured various ecological and physiological profiles of plants in a mercury-contaminated area. In the arid ecosystem, mercury emissions proved dominant in the mercury cycle, while plants functioned as channels for the interphase transfer of mercury from the geosphere to the atmosphere.
Asati et al. also examined the effect of heavy metals, including mercury, on plants and their metabolic activity in areas with high anthropogenic pressure [9]. Heavy metals appeared to have a severe toxic effect on plants, animals, and other local living organisms. Jameer Ahammad et al. claimed that even low concentrations of mercury has a negative effect on plants, e.g. stunted growth and many other adverse consequences [10].
High levels of mercury in soil demonstrated various adverse effects on plant growth and metabolism, e.g. poor photosynthesis, transpiration, water absorption, chlorophyll synthesis, and high lipid peroxidation [11–15].
In plants, a high content of mercury affects most enzymes. Zhou et al. studied the global database for about 35 000 measurements of mercury [16]. They examined the distribution and absorption of mercury in deciduous and coniferous ecosystems. The scientists believe that an effective monitoring of the impact of vegetation on the global mercury cycle requires a better parameterization of models and more consistent observational data, while recording the exchange of mercury in the entire ecosystem is especially important.
Obrist et al. investigated the role of sedimentation in the global cycle of mercury [17, 18]. The precipitation of mercury compounds occurs all year round. However, it is much higher in summer because the metal is absorbed by vegetation. Absorption of gaseous mercury by the tundra increases its concentration in the soil. The authors predict an increase in the impact of mercury on various ecosystems and human life, which requires further multifaceted research.
Ranieri et al. discovered that phytoextraction is an effective and affordable technological solution for the removal of metals, including mercury, from contaminated soil and water [19]. Jiskra et al. confirmed the severe effect of mercury isotopes on mercury absorption by vegetation [20]. Greger et al. studied six plant species that translocate and release mercury into the air [21]. They used a transpiration chamber to monitor the absorption of mercury by the roots, its further distribution over the shoots, and the final release through the shoots. The research featured garden peas, spring wheat, sugar beets, oilseed rape, white clover, and willow. All the plants were able to absorb significant amounts of mercury from its nutrient solution (200 μg/L). However, the translocation to the shoots was rather low (0.17–2.50 %).
Juillerat et al. examined soil and ground litter in 15 locations covered by northern deciduous trees or mixed deciduous and coniferous forests [22]. Their research objective was to determine how mercury content depended on the tree species, forest type, and soil profile. Twelve tree species from two sites demonstrated significant differences. The research proved that the peculiarities of a particular territory are important for mercury studies. The differences in the mercury pools from ground litter correlated with the differences in carbon pools.
These global issues are relevant for Russia and the Kemerovo Region. Komov et al. studied the content of mercury in soil, water sediments, and animals on the banks of the Rybinsk Reservoir [23]. The recorded mercury concentrations varied by more than two orders of magnitude. As for aquatic invertebrates, the concentration of metal appeared to be high in heterotopic species: larvae and adult insects had 0.85 mg of mercury per 1 kg of dry weight. However, homotopic species had a lower concentration of mercury, e.g. for mollusks, it was 0.11 mg per 1 kg of wet weight. As for predatory arachnids, aquatic and semi-aquatic species proved to have higher concentrations of mercury: for hydrocarina, it was ≤ 0.68, and for raft spiders, it was ≤ 0.33 mg per 1 kg of dry weight. On the contrary, spiders that lived far from water sources revealed much lower concentrations of mercury: crab spiders ≤ 0.07 mg per 1 kg of dry weight. Creatures that feed on vegetation or phytophagous animals also demonstrated lower mercury concentrations.
Gremyachikh et al. studied the content of mercury in the muscle tissue of river perch fished in different areas of the Rybinsk Reservoir in 1997–2012 and registered an increase in mercury concentration in recent decades [24].
Gorbunov et al. assessed the increase in mercury in the tissues of fish caught in the Volga [25]. They focused on how the accumulation of mercury in the muscle tissues of perch, bream, and pike depended on the mass of fish. The research registered a directly proportional dependence for perch (correlation coefficient r = 0.881, p = 0.018) and an inversely proportional relationship for pike (r = –0.653, p = 0.029). For bream, no such dependence was revealed.
Komov et al. studied the content of mercury in five species of amphibians and seven species of leeches [26]. The average values for amphibians were 0.007–0.101, for leeches – 0.014–0.065 mg per 1 kg of wet weight. The concentration of mercury depended on the taxonomy, habitat, and tissue type. The experiment established some consequences of the alimentary mercury intake on several biological parameters, i.e. metamorphosis rate of toad larvae, behavior pattern of tadpoles of frogs and leeches, etc. The results delivered new data on the mechanisms of migration and distribution of mercury compounds in aquatic, near-water, and terrestrial ecosystems.
Golovanov et al. studied in vivo the effect of accumulated mercury on the maltase and amylolytic activity of glycosidases in tadpoles of the common toad (Bufobufo L.) [27]. The research revealed changes in the activity of glycosidases depending on the level of accumulated mercury and the timing. The activity of the glycosidases decreased, whereas the sensitivity of starch-hydrolyzing enzymes to heavy metal ions (Cu, Zn, Cd, and Pb) increased.
The physicochemical properties of mercury allow it to circulate, accumulate, and redistribute in environment, depending on the particular conditions of aquatic and terrestrial ecosystems. Most of the mercury is dispersed and creates a natural global geochemical background, superimposed on man-induced mercury pollution, thus forming areas of antropogenic pollution.
Until recently, the accumulation of mercury by hydrobionts attracted most scientific attention because aquatic environment is optimal for the formation of the most toxic organomercury compounds. Methylated mercury compounds accumulate in living organisms more intensively than inorganic ones and are slowly to excrete. As a result, the transport of mercury along the food chain is faster than in cases of direct absorption of the metal from the environment.
The content of mercury in living organisms increases at the tops of food webs and reaches maximal values in predatory fish, fish-eating birds, and mammals.
Terrestrial ecosystems attract less attention regarding the issues of mercury accumulation and distribution. More research is needed to establish the accumulation patterns of mercury compounds by living organisms in terrestrial ecosystems. The best way to establish the patterns is to determine the level of mercury accumulation in organisms of different trophic groups.
The present research objective was to study the mercury accumulation and its effect on various components of terrestrial ecosystems near the Beloosipovo mercury deposit (Kemerovo region, Russia).
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
The research featured such components of the terrestrial ecosystem as soil, herbaceous plants, herpetobiont insects, and small mammals harvested in the vicinity of the Beloosipovo mercury deposit in the Kemerovo region, Russia (55.196730 N, 86.970065 E).
The sampling involved standard methods. Regardless
of the wind pattern, all samples were taken at four
cardinal points (Fig. 1) at three radii:
1) 0.5 km from the pollution source;
2) 1.5 km from the pollution source;
3) 3.0 km from the pollution source.
The sampling points:
Point 0 (Сontrol) – N 55°10.920ꞌ, E 087º00.959ꞌ
Point North 1 (N1) – N 55°11.180ꞌ, E 087°00.980ꞌ
Point North 2 (N2) – N 55°11.798ꞌ, E 087°00.954ꞌ
Point North 3 (N3) – N 55°12.561ꞌ, E 087°01.244ꞌ
Point South 1 (S1) – N 55°10.654ꞌ, E 087°00.958ꞌ
Point South 2 (S2) – N 55°10.189ꞌ, E 087°01.146ꞌ
Point South 3 (S3) – N 55°09.654ꞌ, E 087°01.123ꞌ
Point West 1 (W1) – N 55°10.915ꞌ, E 087°00.605ꞌ
Point West 2 (W2) – N 55°10.918ꞌ, E 086°59.920ꞌ
Point West 3 (W3) – N 55°10.940ꞌ, E 086°58.496ꞌ
Point East 1 (E1) – N 55°10.866ꞌ, E 087°01.333ꞌ
Point East 2 (E2) – N 55°10.939ꞌ, E 087°02.427ꞌ
Point East 3 (E3) – N 55°10.876ꞌ, E 087°03.705ꞌ

The territory of the Beloosipovo mine was considered as the main source of pollution and marked as Point 0 (C).
The control sampling was carried out at N 55°13.291ꞌ, E 086°35.294ꞌ. It was located more than 30 km away from the Beloosipovo mercury deposit, which means it had no effect whatsoever on the background indicators. The soil sampling followed State Standards R 56157-2014 and State Standards 17.4.3.01-2017 using the envelope method at a depth of 0–20 cm and 30–60 cm. The total sampling weight was ≥ 2 kg. The soil samples were put in separate plastic containers and labeled.
The eight herb samples were taken in the same areas as the soil samples. The combined sample wet weight was ≥ 2 kg (natural moisture). The plants were removed together with the rhizomes, which were thoroughly cleared of soil. The samples were placed in plastic bags and labeled.
Invertebrates are the main link by which mercury from the environment enters the organisms of vertebrates. Herpetobiont insects inhabit the soil surface and are widespread in terrestrial ecosystems. They play an important role in food and soil chains.
The main group of herpetobiontic insects was represented by four families of Coleoptera (Coleoptera L.): dung beetles (Geotrupidae L.), lamellar beetles (Scarabaeidae L.), ground beetles (Carabidae L.), and istafilinids (Staphylinidae L.).
The herpetobiont insects were caught using Barber’s traps. At one point, 50 traps with a volume of 0.3 l were dug in one line at a distance of 1 m from each other. The traps contained 5% acetic acid solution. The insects collected at each point were packed into containers, labeled, and stored in an automobile refrigerator at –4°C.
The small mammals were represented by insectivores (Eulipotyphla L.) and rodents (Rodentia L.). They were caught using crushers. At each point, 50 crushers were installed at a distance of 1 m from each other. The captured animals were placed in plastic containers, labeled, and stored in a car refrigerator.
The species composition of the mammals:
Byinsectivores (Eulipotyphla)
Shrews (Soricidae L.):
– Common shrew (Sorex araneus L.);
– Even-toothed shrew (Sorex isodon L.);
– Pygmy shrew (Sorex minutus L.);
– Masked shrew (Sorex caecutiens L.);
– Water shrew (Neomys fodiens L.).
Rodents (Rodentia)
Hamsters (Cricetidae L.):
– Red-backed vole (Clethrionomys rutilus L.);
– Grey-sided vole (Clethrionomys rufocanus L.);
– Bank vole (Clethrionomys glariolus L.);
– Root vole (Microtu oeconomus L.);
– Common field vole (Microtus agrestis L.).
Mice (Muridae L.):
– Field mouse (Apodemus agrarius L.);
– Jerboa mouse (Dipodidae L.);
– Birch mouse (Sicista betulina L.).
The sampling of water in the Belaya Osipova river was carried out 0.5–1.0 km above the mouth (Fig. 2) in five replicates in 2018–2021. The samples were poured into two-liter vessels and were delivered to the laboratory within no more than 18 h from the moment of water intake.
The concentration of mercury in soil, plants, herpetobiont insects, and small mammals was carried out in an accredited laboratory of the Kemerovo State University (Russia). The tests followed Federal Environmental standard PNDF 16.1:2:2.2.80-2013 (М 03-09-2013) “Quantitative chemical analysis of soils. Methods for measuring the mass fraction of total mercury in samples of soils and grounds, including greenhouses, clays, and bottom sediments, by the atomic absorption method using a mercury analyzer RA-915M.”
The samples were prepared using wet mineralization and concentrated nitric acid, hydrochloric acid, and hydrogen peroxide. The samples were dried to obtain biosubstrates in an EKPS-10 electric chamber furnace at 520°C. The obtained white ash was used to determine the content of mercury.
The method for measuring the mass fraction of total mercury involved thermal decomposition accompanied by the atomization of mercury. After that, the atomic mercury was transferred to the analytical cell of the analyzer by air flow. The atomic absorption of mercury was measured at a resonant wavelength of 253.7 nm. The mass fraction of mercury in the sample was automatically determined by the peak area value (analytical signal). The process was based on the preset calibration characteristic using the software for the analyzer (RAPID software). The calibration was carried out using standard samples of a solution of mercury ions. It involved a calibration sample that contained mercury adsorbed on activated carbon.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
According to the e-catalog of geological documents from the Russian Federal Geological Fund, the mercury deposit of 124 tons is located in the river basin of the Belaya Osipova. The ore-bearing mineral is cinnabar (HgS). Mineralization is extremely uneven, and areas of high concentration are replaced by barren ones. The deposit has a hydrothermal low-temperature origin and is confined to the zone of deep and echelon faults. The study area has manifestations and mineralization points, as well as placer and geochemical aureoles of mercury.
The deposit was developed in 1969–1975. A small plant extracted mercury from ore by evaporation. No exact information on the volume of mined mercury is available; according to unofficial data, it mined only several tens of tons.
The area under study is covered by black forests of Siberian fir (Abies sibirica Ledeb.), Aspen (Populus tremula L.), birch (Betula pubescens Ehrh., Betula pendula Roth), and tall grasses, which can reach three meters in height.
The lush undergrowth is represented by such shrubs as goat willow (Salix caprea L.), cranberry bush (Viburnum opulus L.), pea shrub (Caragana arborescens Lam.), Siberian mountain ash (Sorbus sibirica Hedl.), and bird cherry (Padus avium Mill.). Some areas have scarce undergrowth.
The most typical herbaceous plant species are melancholy thistle (Cirsium heterophyllum (L.) Hill.), millet grass (i>Milium effusum< L.), dissected hogweed (Heracleum dissectum Ledeb.), wild chervil (Anthriscus sylvestris (L.) L.), Siberian cacalia (Crepis sibirica L.), northern wolfsbane (Aconitum septentrionale Koelle), black meadowsweet (Filipendula ulmaria (L.) Maxim.), Siberian globeflower (Trollius asiaticus L.), giant fescue (Festuca gigantea (L.) Vill.), etc.
The area has a big population of large ferns, which often dominate the herbaceous cover: adderspit (Pteridium aquilinum (L.) Kuhn.), male shield fern (Dryopteris filix-mas (L.) Schott), female fern (Athyrium filix femina (L.) Roth), and ostrich fern (Matteuccias truthiopteris (L.) Tod.).
Nemoral tertiary relics are represented by alfredia (Alfredia cernua (L.) Cass.), giant fescue (F. gigantea (L.) Vill.), whitespot betony (Stachys sylvatica L.), male shield fern (D. filix-mas (L.) Schott), sweet woodruff (Galium odoratum (L.) Scop.), and slender false brome (Brachypodium sylvaticum (Huds.) Beauv.).
The area is dominated by forest phytocenoses, mostly tall-grass forests with a forest stand of birches, aspens, and firs (2Os3B5P): drooping birch (B. pendula Roth.), Siberian fir (A. sibirica Ledeb.), aspen (Populus tremula L.), and Siberian spruce (Picea obovata Ledeb.). Siberian fir and silver birch have a good seed reproduction; as a result, the forest canopy is rich in fir undergrowth, while the open areas demonstrate a thick population of young birches. The average diameter of the birch is ≤ 40 cm, the average height is 25 m. The average diameter of the fir is ≤ 30–40 cm, the height is 28–30 m. The average diameter of the aspen is 40–50 cm, the height is about 30 m, and the crown density can reach 0.7–0.8.
The composition of the forest stand differs in the ratio of fir, aspen, and birch: birch-fir-aspen, fir-aspen, or aspen-fir with a few birches, while some areas are entirely fir or birch forests. Some areas have a rich undergrowth: goat willow (S. caprea L.), cranberry bush (V. opulus L.), pea shrub (C. arborescens Lam.), red raspberry (Rubusidaeus L.), Siberian mountain ash (S. sibirica Hedl.), d owny c urrant (Ribes spicatum Robson.), black currant (Ribes nigrum L.), and bird cherry (Padusavium Mill.).
In the open and birch-dominated areas, raspberries grow in lush thickets. Some forest parts have a steeplysloping terrain with areas of higher moisture, where willow thickets proliferate. Willow patches and firor aspen-predominated areas also host vines, usually represented by wild hop (Humulus lupulus L.).
The grass stand is represented by tall grasses. The projective cover is over 85%. The maximal height of the grass standcan reach 3.5 m in cases of alfredia or hogweed, while the average height is 1.5 m. The list of tall grasses includes: melancholy thistle (C. heterophyllum (L.) Hill.), millet grass (M. effusum L.), northern wolfsbane (A. septentrionale Koelle), dissected hogweed (H. dissectum Ledeb.), meadow rue (Thalictrum minus L.), golden thoroughwax (Bupleurum aureum Fisch. ex Hoffm.), great nettle (Urtica dioica L.) wild chervil (A. sylvestris (L.) Hoffm.), meadowsweet (F. ulmaria (L.) Maxim.), cacalia (Cacalia hastata L.), and Siberian hawk’s beard (C. sibirica L.). In some places, especially those dominated by fir trees, the thickets are formed almost entirely by nettle, infested by dodder (Cuscuta sp.).
Other perennial herbs also play a significant role in the composition of the phytocenosis: alfredia (A. cernua (L.) Cass.), four-leaved Paris herb (Parisqua drifolia L.), wood geranium (Geranium sylvaticum L.), Dahurian chickweed (Cerastium davuricum Fisch. ex Spreng.), Bunge chickweed (Stellaria bungeana Fenzl.), wood sorrel (Oxalisa cetosella L.), Siberian globeflower (T. asiaticus L.), wild leek (Allium microdictyon Prokh.), lungwort (Pulmonaria mollis Wulf. ex Hornem), spurge (Euphorbia pillosa L.), touch-me-not (Impatiens nolitangere L.), Urals peony (Paeonia anomala L.), northern bedstraw (Galium boreale L.), sedge (Carex macroura Meins.), Greek-valerian polemonium (Polemonium caeruleum L.), violet (Violauni flora L.), whitespot betony (S. sylvatica L .), and snakeflower (Lamium album L.).
Ferns make up part of some grass stand areas: female fern (Athyrium filix-femina (L.) Roth), adderspit (P. aquilinum (L.) Kuhn.), ostrich fern (Matteuccia struthiopteris (L.) Tod.), and male shield fern (D. filixmas (L.) Schott). Adderspit and ostrich grow in thickets.
The herbaceous layer also includes species from the spring synusia, which have completed their growing season (Corydalis, Anemone s. L., etc.), including Siberian trout lily (Erythronium sibiricum (Fisch. et C. A. Mey) Kryl.). This flower is endemic to the Altai- Sayan ecoregion and is protected by the federal and regional law.
Herb-dominated patches appear in some open spaces, depending on the moisture and some other factors. They form tall-grass-grassland patches, grass meadows, and motley grass-grasses associations.
The tall-grass-grassland meadows consist of the same species as the herb layer in the forest: melancholy thistle (C. heterophyllum (L.) Hill.), northern wolfsbane (A. septentrionale Koelle), dissected hogweed (H. dissectum Ledeb.), meadow rue (T. minus L.), golden thoroughwax (Bupleurum aureum Fisch. hastata L.), wild chervil (C. sibirica L.), wild leek (A. microdictyon Prokh.), soft lungwort (P. mollis Wulf. ex Hornem), spurge (E. pillosa L.), etc.
The grass meadows and motley grass-grasses associations develop on sunlit and warm areas, e.g. forest edges. Some species grow both in the forest and in the open, e.g. meadow rue (T. minus L.), golden thoroughwax (B. aureum Fisch. ex Hoffm.), wild chervil (Anthris cussylvestris (L.) Hoffm.), meadowsweet (F. ulmaria (L.) Maxim.), cock’s-foot (Dactylis glomerata L.), bluegrass (Poa sp.), timothy grass (Phleum pratense L.), common tansy (Tanacetum vulgare L.), lousewort (Pedicularis incarnata L.), bladder campion (Oberna behen (L.) Ikonn.), etc. More humid areas are home to other kinds of bluegrass (Poa remota Forsell.), water forget-me-not (Myosotis palustris (L.), white hellebore (Veratrum lobelianum Bernh.), Siberian globeflower (T. asiaticus L.), buttercup (Ranunculus sp.), marsh orchid (Dactylorhiza sp.), wood bulrush (Scirpus sylvaticus L.), clump speedwell (Veronica longifolia L.), groundsel (Senecio sp.), etc.
Many meadows are gradually overgrowing with willow and birch. Willow thickets predominate in the floodplain of the river and represented by goat willow (S. caprea L.), woollytwig willow (Salix dasyclados Wimm.), basket willow (Salixvim inalis L.), almondleaved willow (Salix triandra L.), etc. The list of herbs that proliferate in the willow patches includes fireweed (Chamerion angustifolium (L.) Holub), wood horsetail (Equisetumsyl viaticum L.), common loosestrife (Lysim achiavulgaris L.), sedge (Carex sp.), etc.
In addition to willow thickets, floodplain meadows are also widespread along the river banks, where grain grass prevails, e.g. smallweed (Calamagrostis sp.), cock’s-foot, timothy grass, etc. The floodplain areas also include white hellebore (V. lobelianum Bernh.), sorrel (Rumex sp.), marsh orchid (Dactylorhiza sp.), wood reed (S. sylvaticus L.), clump speedwell (V. longifolia L.), ragged robin (Coccyganthe flos-cuculi (L.), dissected hogweed (H. dissectum Ledeb.), marsh cress (Rorippapalustris (L.) Bess.), lousewort (Scrophularia sp.), scouring horsetail (Equisetum hiemale L.), sedge (Carex sp.), common loosestrife (L. achiavulgaris L.) angelica (Archangelica decurrens Ledeb.), and coltsfoot (Tussilago farfara L.). Angelica grows in lush thickets. Birch and bird cherry also grow on the floodplain meadows.
In shallow water, there are thickets of butterbur (Petasites radiatus (J.F. Gmel.) J. Toman) and rush flower (Butomusum bellatus L.).
Therefore, the study area boasts a significant biological diversity. In addition, it is home to a species protected at the federal and regional levels, namely Siberian trout lily (E. sibiricum (Fisch. Et C. A. Mey) Kryl.) and several nemoral tertiary relics, such as alfredia (A. cernua (L.) Cass.), giant fescue (F. gigantea (L.) Vill.), whitespot betony (S. sylvatica L.), male shield fern (D. filix-mas (L.) Schott), and slender false brome (B. sylvaticum (Huds.) Beauv.). No invasive species were registered.
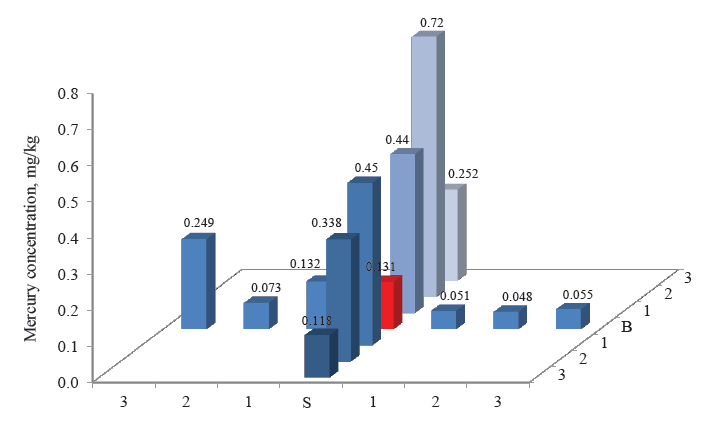
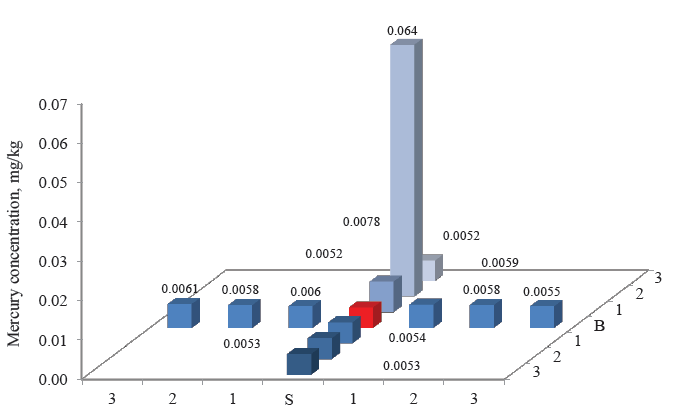
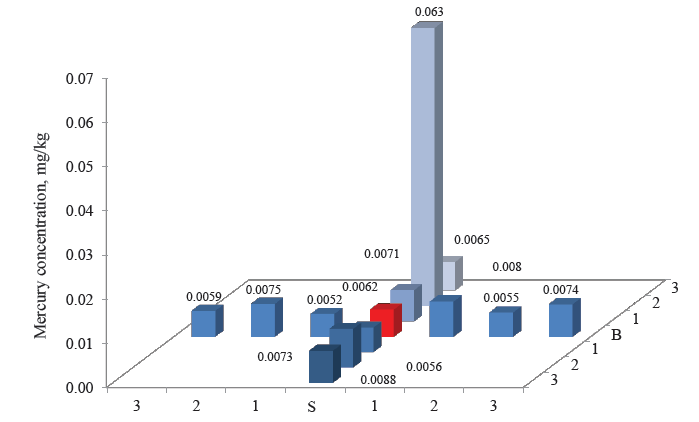
Figures 3–8 demonstrate the mercury concentration in soil, plants, insects, and small mammals near the Beloosipovo mercury deposit and in the control zone. The highest concentration of mercury was observed at point North 2 (N 2), which was located at 1.5 km north of the deposit: in soil – 0.72 mg/kg and 0.96 mg/kg, in plants – 0.064 mg/kg, in insects – 0.063 mg/kg, in rodents – 0.091 mg/kg, and in insectivores – 0.056 mg/kg.

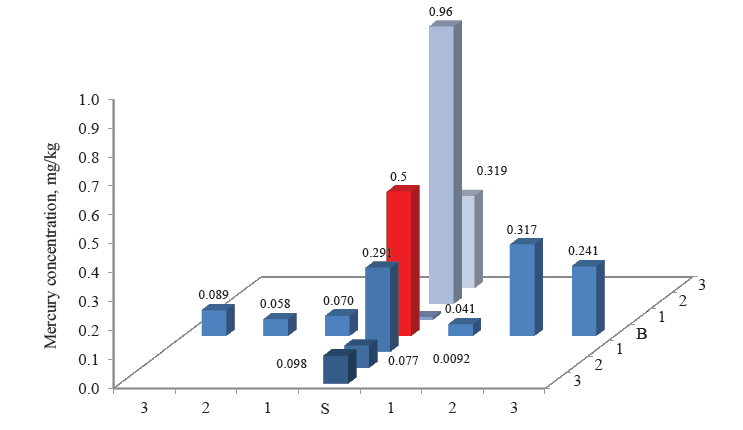
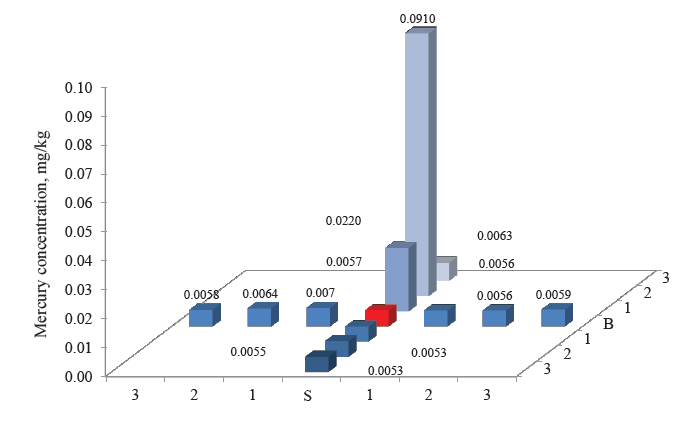

According to regulatory documents, the maximal permissible concentration of mercury in soil is 2.1 mg/kg. As the maximal value in the soil samples was 0.96 mg/kg, it means that no dangerous concentration of mercury was detected. However, the e-catalog of geological documents specifies the average concentration of mercury in the soils of the Kemerovo Region at the level 0.16–0.22 mg/kg [28]. Thus, the concentrations of mercury in the soil near the Beloosipovo mercury deposit proved to be by 3–4 times higher than the average values, despite the fact that the mine was closed more than 40 years ago.
The high concentration of mercury in the samples taken the north was presumably related to the terrain peculiarities: the altitude decreases from north to south, dropping from 407 to 214 m. Points North 2 (N 2) and North 3 (N3) were located directly in the deposit zone, while point North 1 (N 1) was on the borderline.
As for the control point, the concentration of mercury in all components of the ecosystem was much lower than in the area under analysis: in soil and small mammals, it was lower by 1–3 orders of magnitude; in plants and herpetobiontic insects – by 2–4 times.
While the soil samples demonstrated a permissible concentration of mercury, the samples from the Belaya Osipova river exceeded the permissible value (Fig. 9). The maximal permissible concentration of mercury for water bodies is 0.0005 mg/L. In the Belaya Osipova (2018–2021), the concentration exceeded the permissible value by 5–20% and reached 0.00056–0.00074 mg/L.
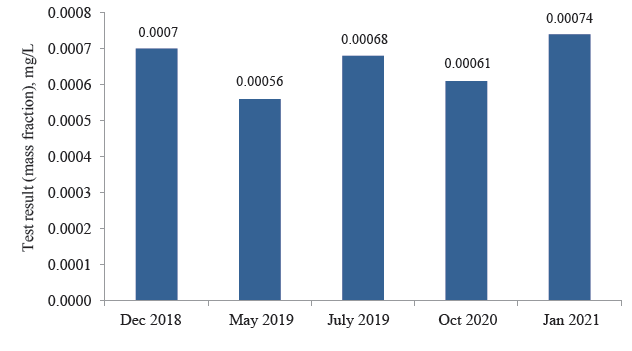
The high content of mercury in the Belaya Osipova may be associated with the Beloosipovo mercury deposit: mercury compounds might be washed out by groundwater and surface spring floods. Further studies require additional tests of the water biocenosis, which will be one of the tasks of subsequent research.
The concentration of heavy metals is believed to increase up the food chains. To test this presumption, we compared the concentration of mercury in the food chains at points North 2 (N 2) and North 3 (N3) with the highest mercury concentration in the soil. However, it was the soil samples that demonstrated the highest concentration of mercury, and further up the food chains its concentration dropped by one or two orders of magnitude, depending on the collection point (Fig. 10). The greatest drop was observed at North 2, where the concentration of mercury in the soil was the highest: from 0.72 to 0.022 mg/kg in the soil – plants – mice chain and from 0.72 to 0.017 mg/kg in the soil – plants – insects – shrews chain.
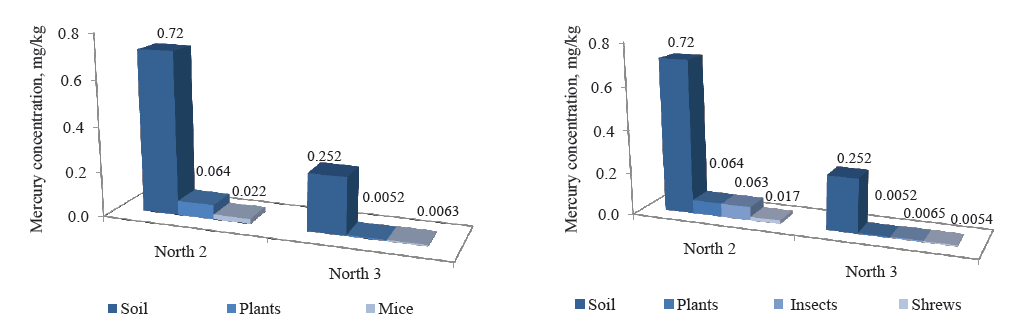
ВЫВОДЫ
The mercury concentration in the soil near the Beloosipovo mercury deposit did not exceed the maximal permissible concentrations. The maximal mercury concentration in the soil was 0.96 mg/kg while the permissible value is 2.1 mg/kg. In the control zone, the research registered a decrease in the mercury concentration by 1–3 orders of magnitude for individual components of the terrestrial ecosystem, namely soil and small mammals. However, the water samples from the Belaya Osipova exceeded the maximal permissible concentration by 5–20% in 2018–2021, which means that mercury compounds may go with groundwater and surface spring floods.
The detected mercury concentrations proved to produce no negative effect on the ecosystem, which was confirmed by the rich biological diversity. The area is home to the critically endangered species of Siberian trout lily (Erythronium sibiricum (Fisch. et C.A. Mey) Kryl.) and several nemoral tertiary relics, such as alfredia (Alfredia cernua (L.) Cass.), giant fescue (Festuca gigantea (L.) Vill.), whitespot betony (Stachys sylvatica L.), male shield fern (Dryopteris filix-mas (L.) Schott), and slender false brome (Brachypodium sylvaticum (Huds.) Beauv.). The research revealed no invasive species.
The mercury content decreased up the food chains, which means that the Beloosipovo mercury deposit has no negative impact on the local ecosystems. The present article is the first part of a series of related publications. Further publications will feature the impact of technogenic centers on the local ecosystem and its individual representatives.
КОНФЛИКТ ИНТЕРЕСОВ
The authors declare no conflict of interests regarding the publication of this article.ФИНАНСИРОВАНИЕ
The research was conducted on the premises of the Research Equipment Sharing Center of Kemerovo State University, agreement No. 075-12021-694 dated August 5, 2021, between the Ministry of Science and Higher Education of the Russian Federation (Minobrnauka) and Kemerovo State University (KemSU) (contract identifier RF----2296.61321X0032).СПИСОК ЛИТЕРАТУРЫ
- Selin H, Keane SE, Wang S, Selin NE, Davis K, Bally D. Linking science and policy to support the implementation of the Minamata Convention on Mercury. Ambio. 2018;47(2):198–215. https://doi.org/10.1007/s13280-017-1003-x.
- Zhou J, Du B, Shang L, Wang Z, Cui H, Fan X, et al. Mercury fluxes, budgets, and pools in forest ecosystems of China: A review. Critical Reviews in Environmental Science and Technology. 2020;50(14):1411–1450. https://doi.org/10.1080/10643389.2019.1661176.
- Karimi E, Yari M, Ghaneialvar H, Kazemi HR, Asadzadeh R, Aidy A, et al. Effects of dust phenomenon on heavy metals in raw milk in western Iran. Foods and Raw Materials. 2020;8(2):241–249. http://doi.org/10.21603/2308-4057-2020-2-241-249.
- Zhu W, Lin C-J, Wang X, Sommar J, Fu X, Feng X. Global observations and modeling of atmosphere-surface exchange of elemental mercury: A critical review. Atmospheric Chemistry and Physics. 2016;16(7):4451–4480. https://doi.org/10.5194/acp-16-4451-2016.
- Gustin MS, Ericksen JA, Schorran DE, Johnson DW, Lindberg SE, Coleman JS. Application of controlled mesocosms for understanding mercury air-soil-plant exchange. Environmental Science and Technology. 2004;38(22):6044–6050. https://doi.org/10.1021/es0487933.
- Fantozzi L, Ferrara R, Dini F, Tamburello L, Pirrone N, Sprovieri F. Study on the reduction of atmospheric mercury emissions from mine waste enriched soils through native grass cover in the Mt. Amiata region of Italy. Environmental Research. 2013;125:69–74. https://doi.org/10.1016/j.envres.2013.02.004.
- Mazur M, Mitchell CPJ, Eckley CS, Eggert SL, Kolka RK, Sebestyen SD, et al. Gaseous mercury fluxes from forest soils in response to forest harvesting intensity: A field manipulation experiment. Science of the Total Environment. 2014;496:678–687. https://doi.org/10.1016/j.scitotenv.2014.06.058.
- Leonard TL, Taylor GE, Gustin MS, Fernandez GCJ. Mercury and plants in contaminated soils: 1. Uptake, partitioning, and emission to the atmosphere. Environmental Toxicology and Chemistry. 1998;17(10):2063–2071. https://doi.org/10.1002/etc.5620171024.
- Asati A, Pichhode M, Nikhil K. Effect of heavy metals on plants: An overview. International Journal of Application or Innovation in Engineering and Management. 2016;5(3):56–66.
- Jameer Ahammad S, Sumithra S, Senthilkumar P. Mercury uptake and translocation by indigenous plants. Rasayan Journal of Chemistry. 2018;11(1):1–12. https://doi.org/10.7324/RJC.2018.1111726.
- Nagajyoti PC, Lee KD, Sreekanth TVM. Heavy metals, occurrence and toxicity for plants: A review. Environmental Chemistry Letters. 2010;8(3):199–216. https://doi.org/10.1007/s10311-010-0297-8.
- Cargnelutti D, Tabaldi LA, Spanevello RM, de Oliveira Jucoski G, Battisti V, Redin M, et al. Mercury toxicity induces oxidative stress in growing cucumber seedlings. Chemosphere. 2006;65(6):999–1006. https://doi.org/10.1016/j.chemosphere.2006.03.037.
- Marrugo-Negrete J, Durango-Hernández J, Pinedo-Hernández J, Enamorado-Montes G, Díez S. Mercury uptake and effects on growth in Jatropha curcas. Journal of Environmental Sciences. 2016;48:120–125. https://doi.org/10.1016/j.jes.2015.10.036.
- Teixeira DC, Lacerda LD, Silva-Filho EV. Foliar mercury content from tropical trees and its correlation with physiological parameters in situ. Environmental Pollution. 2018;242:1050–1057. https://doi.org/10.1016/j.envpol.2018.07.120.
- Fuentes-Gandara F, Herrera-Herrera C, Pinedo-Hernández J, Marrugo-Negrete J, Díez S. Assessment of human health risk associated with methylmercury in the imported fish marketed in the Caribbean. Environmental Research. 2018;165:324-329. https://doi.org/10.1016/j.envres.2018.05.001.
- Zhou J, Obrist D, Dastoor A, Jiskra M, Ryjkov A. Vegetation uptake of mercury and impacts on global cycling. Nature Reviews Earth and Environment.2021;2(4):269–284. https://doi.org/10.1038/s43017-021-00146-y.
- Obrist D, Agnan Y, Jiskra M, Olson CL, Colegrove DP, Hueber J, et al. Tundra uptake of atmospheric elemental mercury drives Arctic mercury pollution. Nature. 2017;547(7662):201–204. https://doi.org/10.1038/nature22997.
- Obrist D, Kirk JL, Zhang L, Sunderland EM, Jiskra M, Selin NE. A review of global environmental mercury processes in response to human and natural perturbations: Changes of emissions, climate, and land use. Ambio. 2018;47(2):116–140. https://doi.org/10.1007/s13280-017-1004-9.
- Ranieri E, Moustakas K, Barbafieri M, Ranieri AC, Herrera-Melián JA, Petrella A, et al. Phytoextraction technologies for mercury- and chromium-contaminated soil: a review. Journal of Chemical Technology and Biotechnology. 2020;95(2):317–327. https://doi.org/10.1002/jctb.6008.
- Jiskra M, E. Sonke J, Agnan Y, Helmig D, Obrist D. Insights from mercury stable isotopes on terrestrial–atmosphere exchange of Hg(0) in the Arctic tundra. Biogeosciences. 2019;16(20):4051–4064. https://doi.org/10.5194/bg-16-4051-2019.
- Greger M, Wang Y, Neuschütz C. Absence of Hg transpiration by shoot after Hg uptake by roots of six terrestrial plant species. Environmental Pollution. 2005;134(2):201–208. https://doi.org/10.1016/j.envpol.2004.08.007.
- Juillerat JI, Ross DS, Bank MS. Mercury in litterfall and upper soil horizons in forested ecosystems in Vermont, USA. Environmental Toxicology and Chemistry. 2012;31(8):1720–1729. https://doi.org/10.1002/etc.1896.
- Komov VT, Gremyachikh VA, Udodenko YuG, Shchedrova YeV, Yelizarov MYe. Mercury in abiotic and biotic components of aquatic and terrestrial ecosystems in the urban settlement on the shore of the Rybinsk Reservoir. Transactions of Papanin Institute for Biology of Inland Waters RAS. 2017;77(80):34–56. (In Russ.). https://doi.org/10.24411/0320-3557-2017-10003.
- Gremyachikh VA, Lozhkina RA, Komov VT. Spatial-temporal variability of mercury content in the river perch Perca fluviatilis Linnaeus, 1758 (Perciformes: Percidae) of the Rybinsk Reservoir at the turn of the XX–XXI centuries. Ecosystem Transformation. 2019;2(2):85–95. (In Russ.). https://doi.org/10.23859/estr-180816.
- Gorbunov AV, Lyapunov SM, Okina OI, Sheshukov VS. Bioaccumulation of mercury in tissues of freshwater fish. Human Ecology. 2018;(11):23–31. (In Russ.). https://doi.org/10.33396/1728-0869-2018-11-26-31.
- Komov VT, Ivanova ES, Gremyachikh VA, Lapkina LN, Kozlova LV, Zheletok EN, et al. The mercury content in the organism of amphibians and leeches from waterbodies of Vologda and Yaroslavl oblasts and experimental verification of its biological consequences. Transactions of Papanin Institute for Biology of Inland Waters RAS. 2017;77(80):57–76. (In Russ.). https://doi.org/10.24411/0320-3557-2017-10004.
- Golovanova IL, Filippov AA, Komov VT, Urvantseva GA, Evdokimov EG. Effect of mercury accumulation on the activity of glycosidase and their sensitivity to heavy metals in toad tadpoles. Transactions of Papanin Institute for Biology of Inland Waters RAS. 2015;72(75):60–65. (In Russ.). https://doi.org/10.24411/0320-3557-2015-10012.
- Ehlektronnyy katalog geologicheskikh dokumentov [Electronic catalog of geological documents] [Internet]. [cited 2018 Jan 18]. Available from: https://rfgf.ru/catalog/index.php.
- Doklad o sostoyanii i okhrane okruzhayushchey sredy Kemerovskoy oblasti v 2018 godu [Report on the state and protection of the environment in the Kemerovo region in 2018] [Internet]. [cited 2018 Jan 18]. Available from: https://www.ecoindustry.ru/gosdoklad/view/523.html.
