

Аннотация
Abiotic and biotic stresses have a major impact on crop growth. Stress affects the root system and decreases the amount of nutrients in fruits. Modern agricultural technologies help replace mineral fertilizers with new generation biopreparation. Unlike chemical fertilizers, biofertilizers reduce the risk of adverse environmental impacts. Of special interest are extremophilic microorganisms able to survive in extreme conditions. We aimed to study the phytostimulating ability of extremophilic bacteria isolated from disturbed lands in the coal-mining region.We isolated microorganisms from disturbed lands and studied their cultural, morphological, and biochemical properties. Then, we determined their ability to synthesize indole-3-acetic acids. The extremophilic bacteria were identified and subjected to biocompatibility testing by co-cultivation. Next, we created consortia of pure cultures and analyzed biomass growth. Finally, the biopreparation was experimentally tested on Trifolium prantense L. seeds.
We isolated 10 strains of microorganisms that synthesized 4.39 to 16.32 mg/mL of indole-3-acetic acid. The largest amounts of the acid were produced by Pantoea spp., Enterococcus faecium, Leclercia spp., Rothia endophytica, and Klebsiella oxytoca. A consortium of Pantoea spp., E. faecium, and R. endophytica at a ratio of 1:1:1 produced the largest amount of indole-3-acetic acid (15.59 mg/mL) and accumulated maximum biomass. The addition of 0.2% L-tryptophan to the nutrient medium increased the amount of indole-3-acetic acid to 18.45 mg/mL. When the T. prantense L. seeds were soaked in the biopreparation (consortium’s culture fluid) at a concentration of 2.5, the sprouts were 1.4 times longer on the 10th day of growth, compared to the control.
The consortium of Pantoea spp., E. faecium, and R. endophytica (1:1:1) stimulated the growth of T. prantense L. seeds. Our findings can be further used to develop biofertilizers for agriculture.
Ключевые слова
Microbial consortium, biopreparation, soil, extremophilic microorganisms, seed germination, Trifolium prantense L.ВВЕДЕНИЕ
By 2050, the world’s population is expected to reach 9.8 billion people [1]. This population growth, as well as industrialization and a decline in agricultural areas, puts enormous pressure on the decreasing supply of food raw materials [2–4]. To solve this problem, agriculture needs to be modernized [5]. Moreover, there is a shortage of macro- and micronutrients in food products. This problem has been caused by environmental pollution, which depletes nutrients in the soil and, consequently, in agricultural crops.
Abiotic and biotic factors have a significant impact on the quality, growth, and yield of agricultural crops [6, 7]. Abiotic effects include drought, salinity, and pollution (e.g., heavy metals or pesticides) [8–10]. Every year, they cause significant economic losses during plant growth. Due to stress, the roots do not absorb the sufficient amount of minerals from the soil, which leads to a lack of nutrients necessary for normal plant growth.
To increase yields, advanced technologies are used in soil cultivation, including land reclamation (drainage and irrigation) and chemical fertilizers. On the one hand, these fertilizers have a positive effect on plant growth and development [11]. Yet, they can also lead to soil acidification, as well as air and water pollution. Moreover, their excessive use worsens the quality of food [12]. For example, Potetnya et al. studied nitrogen and complex fertilizers. They found that nitrogen fertilizers have the greatest effect on plant growth by affecting plant proteins and chlorophyll. Complex fertilizers contain phosphorus and potassium. Although phosphorus is an important biogenic element, it can sometimes provoke slow plant growth and stem development, as well as pigmentation disorders (dirty green or reddish color). Potassium can also have negative effects, causing a lack of water at the cellular level, as well as wilting and twisting of leaves. Therefore, using complex fertilizers is not a universal solution, since the need for nitrogen, phosphorus, and potassium varies from crop to crop [13].
Biofertilizers are considered quite promising fertilizers [14, 15]. They are substances containing living organisms and/or their metabolites. When applied to seeds, plant surfaces, or soil, they colonize the rhizosphere, which promotes growth and availability of primary nutrients [16]. The use of biofertilizers is on the rise, especially in sustainable agriculture and organic farming. As well as being cost-effective, they improve soil fertility, increase plant growth and yield, and make plants more stress-resistant. In addition, biofertilizers help reduce the burden on the environment and improve soil quality [17]. Kha et al., who treated beans with the culture fluid of the bacterial strain Rhizobium tropici, reported its positive effect on the formation of tubercles [7].
Microbial biostimulators are an effective tool for sustainable agriculture. They can increase the yield and quality of agricultural crops by activating physiological and molecular processes in the plants [18]. The phytohormones produced by microbial biostimulators are an effective alternative to the phytohormones industrially produced by chemical synthesis. These substances regulate the physiological processes in plants, including their growth, development, release of nutrients, and adaptation to the environment [19].
Modern agricultural technologies help replace mineral fertilizers with new generation biological products. Of particular importance are native microbiota adapted to specific climatic conditions [20, 21].
Extremophilic microorganisms (extremophiles) are bacteria that can survive under extreme environmental conditions, such as high or low temperatures, lack of oxygen, adaptation to acidic or basic pH values, high salt concentrations, etc. [22]. As a rule, extremophiles are highly resistant to a number of environmental factors, which makes them resistant to specific climatic conditions [23].
In this study, we used consortia of extremophilic microorganisms, which were isolated from the disturbed lands of Kuzbass coal mines, as biofertilizers, or growth phytostimulants. Our aim was to study the ability of these extremophilic bacteria to exhibit phytostimulating properties.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Extremophilic microorganisms were isolated from the surface layer of the technozem in Kuzbass (Kemerovo Oblast). The sampling was performed in the northwestern part of the Prokopievsk-Kiselevsk geological region of Western Siberia (54°14″ north latitude, 86°26″ east longitude). The soil in the study area is represented by overburden rocks of sandy loamy granulometric composition, lacking a fertile layer.
The soils were sampled at a depth of 0–10 cm according to State Standard 17.4.4.02-2017.
The soils in Kuzbass have acidic and slightly alkaline pH values [24, 25]. In addition, the region has a continental or sharply continental climate with low average annual temperatures [26]. Therefore, to isolate extremophilic microorganisms characteristic of this region, soil samples were cultivated under unfavorable conditions according to Zenov et al. [27]. For this, 1 g of soil was added to 5 mL of meat-peptone broth (pH 5.3 and 9.0) consisting of 10 g of meat peptone (LenReaktiv, Russia), 11 g of meat extract (ChemExpress, Russia), 5 g of sodium chloride (ChemExpress, Russia), and up to 1 L of distilled water. Microorganisms were cultivated in an LSI-3016A/LSI-3016R shaker-incubator (Daihan Labtech, South Korea) at 15°C, 100 rpm for 24 h. Further, suspensions were prepared from the subculture diluted to 10–10, 10–11, and 10–12. They were then reinoculated in meat-peptone agar in Petri dishes by the pour plate method at pH 5.3 and 9.0 and cultivated at 15°C for 24 h. The agar consisted of 10 g of dry peptone, 11 g of meat extract, 5 g of sodium chloride, 20 g of agar-agar (LenReaktiv, Russia), and up to 1 L of distilled water. The reinoculation was repeated three times to obtain pure cultures using the streak plate method [28, 29].
To determine cultural properties of the isolated strains, we prepared suspensions of pure cultures at low concentrations, followed by deep plating on meat-peptone agar. Microorganisms were cultivated in a TCO-1/80 SPU thermostat (Smolensk SKTB SPU, Russia) at 25°C for 24 h. Their morphological features were determined by microscopy of a stained fixed smear with methylene blue [30]. The cultures were classified into Gram-positive or Gram-negative according to Belkin et al. [31]. Sporulation was determined according to Pereira et al. and mobility, according to Molofeyeva et al. [32, 33].
Promising strains with phytostimulating activity were selected based on their ability to synthesize indole-3-acetic acid. Indole-3-acetic acid is a phytohormone that stimulates the proliferation of plant cells and enhances the absorption of minerals and nutrients from the soil [21, 34]. To determine its amount, a suspension of microorganisms in meat-peptone broth containing 0.1% L-tryptophan (ChemExpress, Russia) was cultivated for 48 h at 25°C and then centrifuged for 5 min at 10 000 rpm. After that, 1 mL of the supernatant was added to 1 mL of the Salkowski reagent and incubated for 30 min at room temperature. Next, the optical density was measured on a PE-5Z00VI spectrophotometer (UNITEK, Russia) at 535 nm. Based on the results, a calibration graph of the standard solution was created to determine the amount of indole-3-acetic acid [35].
The isolated bacteria were identified using a Vitek 2 Compact automatic microbiological analyzer (BioMerieux, France) with ID-GP and ID-GN cards for Gram-positive and Gram-negative microorganisms, respectively. The cultures were grown on Columbia agar with blood for 48 h at 25°C. Then, a suspension of strains was prepared with a McFarland density of 2.70– 3.30 using a Densichek plus densitometer (BioMerieux, France) [36].
Biocompatibility was determined by co-cultivation. For this, pure cultures were grown on meat-peptone broth for 48 h at 25°C. Then, they were centrifuged for 5 min at 5000 rpm. The test culture was applied to Petri dishes with meat-peptone agar, and the supernatant was added to the wells. Microorganisms were cultivated for 24 h at 25°C [5].
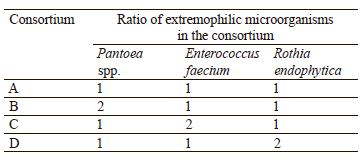
To create consortia, suspensions of pure cultures were prepared in physiological saline with a McFarland density of 0.8–1.1. Then, 5% of the medium containing consortium microorganisms was added to the meatpeptone broth. The resulting consortia of extremophilic bacteria are shown in Table 1.
The amount of indole-3-acetic acid synthesized by the consortia was determined by the method described above.
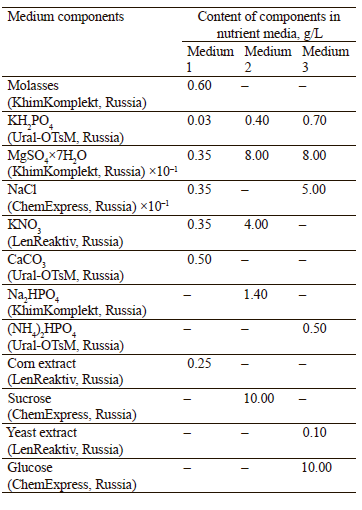
To select the optimal medium composition for the highest biomass yield, microorganisms were cultivated on different media at 25°C for 48 h (Table 2).
Biomass growth was measured according to Zandanova and Gogoleva [37].
To maximize the yield of indole-3-acetic acid, L-tryptophan was added to the previously selected nutrient media in the amount of 0.1, 0.2, and 0.5% of the medium. Microorganisms were cultivated at 25°C for 24 h. The amount of indole-3-acetic acid was measured by the method described earlier.
Red clover (Trifolium pretense L.) is a fodder plant widespread in Kemerovo Oblast. It is a promising plant model for assessing the destructive activity of extremophilic microorganisms against heavy metals and organic contaminants, as well as their phytostimulatory activity [38]. T. pratense L. seeds were prepared in two ways: 1) by soaking in 5 mL of a consortium of microorganisms at a McFarland density of 1.5 and 2.5 for 24 h at 5–8°C; 2) by soaking in 5 mL of distilled water for 24 h at 5–8°C.
Ten seeds were sown into the soil at a depth of 1 cm. They were germinated for 10 days at 18–25°C and relative humidity of 80%. The seeds soaked in the consortia were irrigated with distilled water, while those soaked in distilled water were irrigated with consortia solutions.
Statistical processing was carried out using the Statistica for Windows v. 12.0, (StatSoft, Inc.) at a statistical significance of P = 0 .95. T he t ables a nd fi gures show the arithmetic mean values of the indicators under study. All the experiments were carried out in triplicate.
The equipment for the study was provided by the Instrumental Methods of Analysis in Applied Biotechnology Center at Kemerovo State University.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
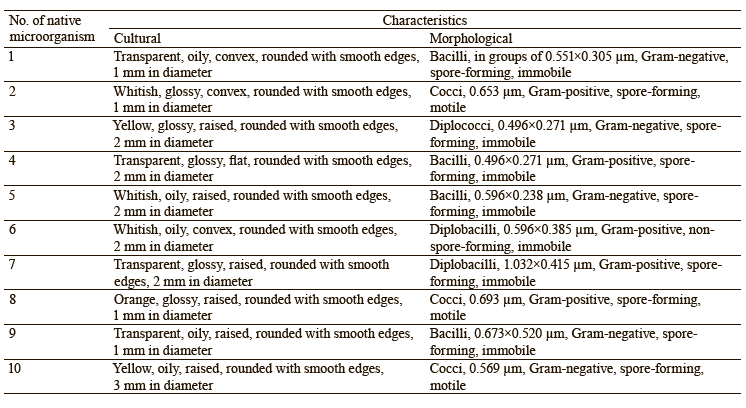
We isolated 10 pure cultures from the soil samples with unfavorable cultivation conditions (acidic and alkaline pH, low temperature). Their cultural and morphological characteristics are presented in Table 3.
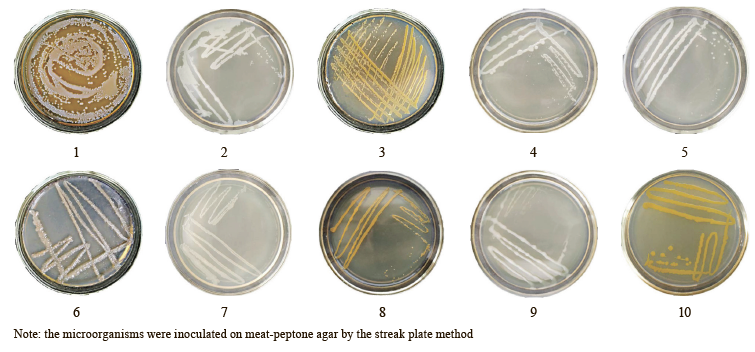
Figure 1 shows the growth of the microorganisms in Petri dishes.
The microscopy results for the isolated strains are shown in Fig. 2.
Table 4 shows the amounts of indole-3-acetic acid synthesized by the isolated microorganisms.

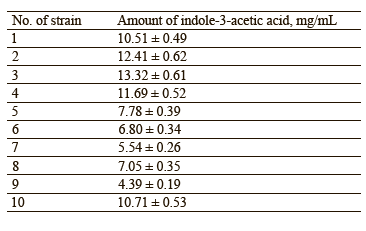
As can be seen from Table 4, the amounts of synthesized indole-3-acetic acid varied from 4.39 to 13.32 mg/mL. The highest and the lowest efficiency was exhibited by strains No. 3 (13.32 mg/mL) and No. 9 (6.39 mg/mL), respectively. As a result, we selected 5 promising strains (No. 1, 2, 3, 4, and 10), which synthesized > 10 mg/mL of indole-3-acetic acid.
To identify the selected isolates, we studied their physical and biochemical characteristics. According to the results, strain No. 1 was Pantoea spp. (probability 0.98), No. 2 was Enterococcus faecium (probability 0.88), No. 3 was Leclercia spp. (probability 0.88), No. 4 was Rothia endophytica (probability 0.86), and No. 10 was Klebsiella oxytoca (probability 0.89). Their biochemical characteristics are presented in Table 5.
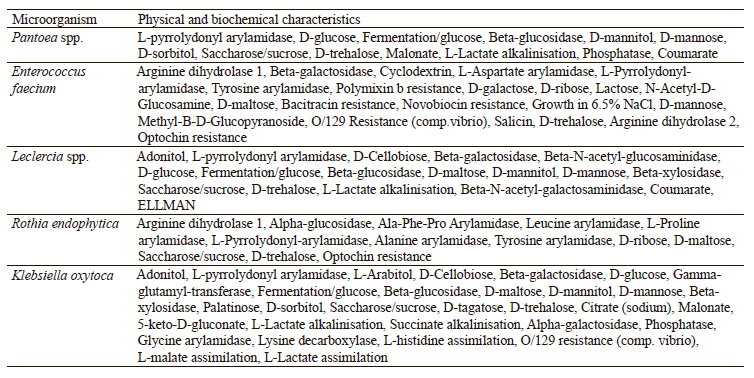
According to Luziatelli et al., the Pantoea species includes agronomically significant strains that are able to stimulate plant growth [39]. In their study, Pantoea strains accumulated 0.12 mg/mL of indole-3- acetic acid. In another study [40], Lee et al. analyzed the effect of E. faecium on plant growth. The authors found that this strain enhanced plant growth and produced 3.63 mg/mL of indole-3-acetic acid. Snak et al. reported that the Leclercia strain stimulated plant growth and synthesized 4.8 mg/mL of indole-3-acetic acid [41]. The study by Shurygin et al. proved that the bacteria of the genus Rothia produced 5.4 mg/mL of indole-3-acetic acid [42]. According to Poveda and Gonzalez-Andres, K. oxytoca showed the maximum yield of indole-3-acetic acid (17.4 mg/mL) [14]. Our results contradicted these published data. This may be due to the fact that we isolated the microorganisms from the Siberian soils and therefore they had greater resistance to adverse environmental conditions and a better ability to produce metabolites.
To create consortia, the microorganisms were tested for biocompatibility (Table 6).

As we can see in Table 6, Leclercia spp. inhibited the growth of Pantoea spp., but, at the same time, exhibited active joint growth with R. endophytica, E. faecium, and K. oxytoca. The K. oxytoca strain was biocompatible only with E. faecium and Pantoea spp., inhibiting the growth of the other strains. Pantoea spp. showed active growth together with E. faecium and R. endophytica, and no growth with Leclercia spp. R. endophytica and E. faecium were biocompatible with all the strains under study. Based on these results, we compiled 4 consortia from Pantoea spp., E. faecium, and R. endophytica.
According to Table 7, the largest amount of indole-3-acetic acid was synthesized by consortium A (15.59 mg/mL). For consortia B, C, and D, the results did not differ significantly. We found that the consortia produced on average 1.4 times more indole-3-acetic acid that the individual microorganisms included in the consortia and about 1.2 times more than Leclercia spp. Thus, the optimal ratio of Pantoea spp., E. faecium, and R. endophytica was 1:1:1.

Then, we selected the optimal composition of the nutrient medium for the highest yield of microbial biomass (Fig. 3). The compositions of the nutrient media are presented in Table 2.
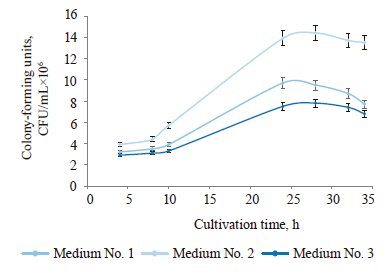
In the first hours of cultivation on media No. 1, 2, and 3, the numbers of colony-forming units were 3.2, 3.9, and 2.9 CFU/mL×106, respectively. The largest number was observed on medium No. 1 after 24 h (9.7 CFU/mL×106), and on media No. 2 and 3 after 28 h (14.4 and 7.8 CFU/mL×106, respectively). Thus, the phase of death began earlier on medium No. 1 than on media No. 2 and 3. However, the number of colonyforming units on medium No. 3 was approximately 2 times lower than on medium No. 2. Thus, we recommend medium No. 2 as optimal for accumulating the largest amount of biomass.
Jahn et al. reported that L-tryptophan is a precursor of indole-3-acetic acid [43]. Therefore, to increase the yield of this phytohormone, we had to select an optimal amount of L-tryptophan. For this, 0.1, 0.2, 0.5% of L-tryptophan was added to medium No. 2. Then, we analyzed biomass growth and the amounts of indole-3- acetic acid synthesized (Table 8).
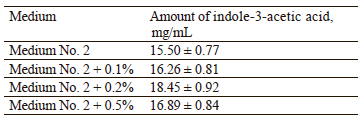
According to Table 8, 0.2% of the volume of medium No. 2 was the optimal amount of L-tryptophan to synthesize 18.45 of mg/mL of indole-3-acetic acid. Thus, we recommended the following composition of the nutrient medium: 0.4 g KH2PO4, 0.8 g MgSO4×7Н2О, 4 g KNO3, 1.4 g Na2HPO4, 10 g sucrose, 2.1 g L-tryptophan, and up to 1 liter of distilled water.
We used Trifolium pretense L. to experimentally evaluate the ability of this biopreparation to enhance plant growth (Table 9).
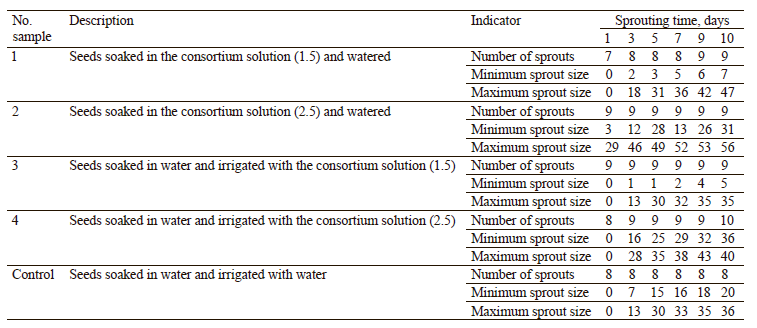
As can be seen from Table 9, samples 1, 2, and 3 had 9 sprouts on the 10th day, while sample 4 had 10 sprouts. Sample 2 had the largest sprout size on the 10th day (56 mm) and sample 3 had the minimum size (5 mm). Figure 4 presents the growth of sprouts during the sprouting time by average length.
On average, the sprouts germinated from the seeds soaked in the consortium solution (2.5) were 6 mm longer than the control on the 3rd day. On the 7th and 10th days, their growth was still effective, with an average length being 14 mm and 1.4 times longer, respectively, than the control. However, when irrigated with the consortium in a lower concentration (1.5), the sprouts were 4 mm shorter than the control. Their growth was less effective and the average length was 7 mm shorter than the control. The minimum length was 1.4 times shorter than that of the control. When the sprouts were irrigated with a higher concentration of the consortium (2.5), they were by 13 mm longer than the control on the 3rd day and 1.3 times as long on the 10th day.
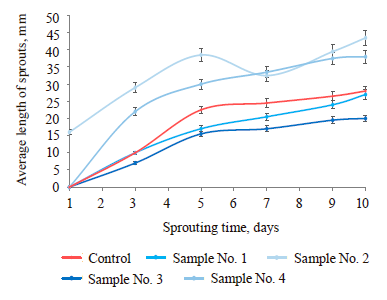
As can be seen from Fig. 5, when soaked in the consortium with a concentration of 2.5, 10 seeds of T. pretense L. developed a root, which is 2 seeds more than the number of seeds soaked in the consortium with a concentration of 1.5. It should be noted that the seeds soaked in water did not develop a root.
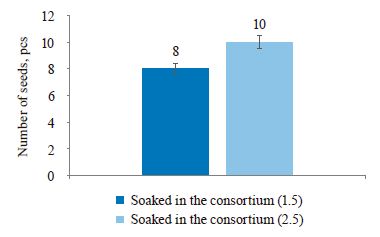
According to Fig. 6, only 2 of the sprouts soaked in the 1.5 consortium had a second cotyledon on the 10th day, compared to 7 sprouts of the control sample. Thus, the consortium with a concentration of 2.5 proved most effective to soak the seeds in, since almost all the seeds (8 out of 10) developed a second cotyledon.
ВЫВОДЫ
We isolated 10 strains of on the amounts of indole-3-acetic acid they synthesized, the most promising strains for consortia were No. 1 (10.51 mg/mL), No. 2 (12.41 mg/mL), No. 3 (16.32 mg/mL), No. 4 (11.69 mg/mL), and No. 10 (10.71 mg/mL). The selected extremophilic bacteria were then identified as Pantoea spp. (No.1), Enterococcus faecium (No. 2), Leclercia spp. (No. 3), Rothia endophytica (No. 4), and Klebsiella oxytoca (No. 10). Based on biocompatibility testing, we created four variants of consortia containing Pantoea spp., E. faecium, and R. endophytica. The largest amount of indole-3-acetic acid was synthesized by a consortium with a 1:1:1 ratio of these extremophilic bacteria (15.59 mg/mL).
The most optimal nutrient medium for biomass accumulation contained 0.4 g KH2PO4, 0.8 g MgSO4×7Н2О, 4 g KNO3, 1.4 g Na2HPO4, 10 g sucrose, and up to 1 liter of distilled water. After 4 h of cultivation, the number of colony-forming units on this medium was 3.9 CFU/mL×106, and the stage of death began after 28 h (14.4 CFU/mL×106).
To increase the content of indole-3-acetic acid, L-tryptophan was added to the nutrient medium in an amount of 0.1, 0.2, and 0.5%. We found that with 0.2% L-tryptophan, the biopreparation synthesized 1.2 times more indole-3-acetic acid than the control.
Trifolium prantense L. was used to assess the ability of the consortium’s culture fluid to enhance plant growth. In the first experiment, the seeds were soaked in the consortium solution at concentrations of 1.5 and 2.5. In the second experiment, the seeds soaked in water were irrigated with the consortium solution at concentrations of 1.5 and 2.5. According to the results, the largest number of sprouts (9 sprouts) was formed from the seeds soaked in the 2.5 consortium solution and from the seeds irrigated with the 1.5 solution.
As for sprout length, the seeds soaked in the biopreparation at a concentration of 2.5 proved the most effective on the 10th day (the sprouts were 1.4 times longer compared to the control). Yet, the sprouts germinated from the seeds irrigated with the biopreparation at the same concentration were 1.3 times shorter compared to the control. We can conclude that soaking is the most effective way to treat seeds.
Thus, plant growth was enhanced by soaking the seeds in the culture fluid of the consortium at a concentration of 2.5 made from Pantoea spp., E. faecium, and R. endophytica (1:1:1) cultivated on medium No. 2 (0.4 g KH2PO4, 0.8 g MgSO4×7Н2О, 4 g KNO3, 1.4 g Na2HPO4, 10 g sucrose, 2.1 g L-tryptophan, and 1000 mL distilled water). Further research will focus on developing biofertilizers to increase the yield and nutritional value of crops.
Вклад авторов
The authors were equally involved in writing the manuscript and are equally responsible for plagiarism.КОНФЛИКТ ИНТЕРЕСОВ
The authors declare no conflict of interest related to this publication.ФИНАНСИРОВАНИЕ
The study was part of the national project “Science and Universities”.СПИСОК ЛИТЕРАТУРЫ
- Sijpestijn GF, Wezel A, Chriki S. Can agroecology help in meeting our 2050 protein requirements? Livestock Science. 2022;256. https://doi.org/10.1016/j.livsci.2022.104822
- Krivikhina AA. Analysis of demographic situation in different countries of the world. SCIFF. Questions of Students Science. 2019;39(11):23–28. (In Russ.).
- Reshetnik EI, Gribanova SL, Egorov DV, Gritsov NV. Plant materials use in the production of fermented milk products for specialized nutrition. Food Industry. 2021;6(4):39–46. (In Russ.). https://doi.org/10.29141/2500-1922-2021-6-4-4
- Rada AO, Kuznetsov AD. Digital inventory of agricultural land plots in the Kemerovo Region. Foods and Raw Materials. 2022;10(2):206–215. https://doi.org/10.21603/2308-4057-2022-2-529
- Volkova GS, Kuksova EV, Serba EM. Investigation of biological interstrains and growing properties of lactic acid bacteria production strains. Topical issues of the dairy industry, intersectoral technologies and quality management systems. 2020;1(1):104–109. (In Russ.). https://doi.org/10.37442/978-5-6043854-1-8-2020-1-104-109
- Jasim B, Jimtha JC, Shimil V, Jyothis M, Radhakrishan EK. Studies on the factors modulating indole-3-acetic acid production in endophytic bacterial isolates from Piper nigrum and molecular analysis of ipdc gene. Journal of Applied Microbiology. 2014;117(3):786–799. https://doi.org/10.1111/jam.12569
- Kha TZ, Kanarskii AV, Kanarskaia ZA, Shcherbakov AV, Shcherbakova EN. The key plant growth stimulator – rhizobacteria. Vesting of Volga State University of Technology. Series: Forest. Ecology. Nature Managemen. 2020;47(3):58–73. (In Russ.). https://doi.org/10.25686/2306-2827.2020.3.58
- Fotina NV, Emelianenko VP, Vorob’eva EE, Burova NV, Ostapova EV. Contemporary biological methods of mine reclamation in the Kemerovo Region – Kuzbass. Food Processing: Techniques and Technology. 2021;51(4):869–882. (In Russ.). https://doi.org/10.21603/2074-9414-2021-4-869-882
- Milentyeva IS, Fotina NV, Zharko MYu, Proskuryakova LA. Microbial treatment and oxidative stress in agricultural plants. Food Processing: Techniques and Technology. 2022;52(4):750–761. (In Russ.). https://doi.org/10.21603/2074-9414-2022-4-2403
- Onipchenko VG, Kazantseva ЕS, Elumeeva TG, Zakharova ЕА, Petrova SЕ, Akhmetzhanova AA, et al. Abiotic factors have stronger effect on functional traits than biotic ones: The transplantation experiments in alpine belt. Journal of General Biology. 2020;81(5)352–361. (In Russ.). https://doi.org/10.31857/S0044459620050097
- Nazaryuk VM, Kalimullina FR. Effect of fertilizers and plant residues on the fertility of soil and the yield and chemical composition of grain crops. Agrohimia. 2010;(6):18–27. (In Russ.).
- Zhang R, Li Y, Zhao X, Degen AA, Lian J, Liu X, et al. Fertilizers have a greater impact on the soil bacterial community than on the fungal community in a sandy farmland ecosystem, Inner Mongolia. Ecological Indicators. 2022;140. https://doi.org/10.1016/j.ecolind.2022.108972
- Potetnya KM, Sadov AA, Virova OM, Pankov YuV. Role and types of fertilizers in agriculture. Scientific and Technical Bulletin: Technical Systems in Agriculture. 2019;5(5):25–32. (In Russ.).
- Poveda J, Gonzalez-Andres F. Bacillus as a source of phytohormones for use in agriculture. Applied Microbiology and Biotechnology. 2021;105:8629–8645. https://doi.org/10.1007/s00253-021-11492-8
- Sezgin M, Kahya M. Phytohormones. Bitlis Eren University Journal of Science and Technology. 2018;8(1):35–39. https://doi.org/10.17678/beuscitech.386726
- Ramírez-Cariño HF, Morales I, Guadarrama-Mendoza PC, González-Terreros E, Martínez-Gutiérrez GA, Dunlap CA, et al. Biofertilizing effect of putative plant growth promoting rhizobacteria in vitro and in tomatillo seedlings (Physalis ixocarpa Brot.). Scientia Horticulturae. 2023;308. https://doi.org/10.1016/j.scienta.2022.111567
- Brahmaprakash GP, Sahu PK. Biofertilizers for sustainability. Journal of the Indian Institute of Science. 2012;92:37–62.
- Kerchev P, van der Meer T, Sujeeth N, Verlee A, Stevens CV, Breusegem FV, et al. Molecular priming as an approach to induce tolerance against abiotic and oxidative stresses in crop plants. Biotechnology Advances. 2020;40. https://doi.org/10.1016/j.biotechadv.2019.107503
- Kumaraswamy RV, Kumari S, Choudhary RC, Pal A, Raliya R, Biswas P, et al. Engineered chitosan based nanomaterials: Bioactivities, mechanisms and perspectives in plant protection and growth. International Journal of Biological Macromolecules. 2018;113:494–506. https://doi.org/10.1016/j.ijbiomac.2018.02.130
- Drozdova MYu, Pozdnyakova AV, Osintseva MA, Burova NV, Minina VI. The microorganism-plant system for remediation of soil exposed to coal mining. Foods and Raw Materials. 2021;9(2):406–418. https://doi.org/10.21603/2308-4057-2021-2-406-418
- Zaghian S, Shokri D, Emtiazi G. Co-production of a UV-stable bacteriocin-like inhibitory substance (BLIS) and indole-3-acetic acid hormone (IAA) and their optimization by Taguchi design in Bacillus pumilus. Annals of Microbiology. 2012;62:1189–1197. https://doi.org/10.1007/s13213-011-0359-6
- Krivushina AA, Startsev VO. Micromycetes-destructors of polymeric materials among extremophilic microorganisms (review). Proceedings of VIAM. 2022;107(1):123–134. (In Russ.). https://doi.org/10.18577/2307-6046-2022-0-1-123-134
- Morozkina EV, Slutskaya ES, Fedorova, TV, Tugay TI, Golubeva LI, Koroleva OV. Extremophilic microorganisms: Biochemical adaptation and biotechnological application (review). Applied Biochemistry and Microbiology. 2010;46(1):1–14. https://doi.org/10.1134/S0003683810010011
- Stepanova OI, Prosyannikov VI. Acidity of arable soils and efficiency of liming in the Kemerovo region. Agrochemical Herald. 2019;(3):37–41. (In Russ.).
- Edelev AV, Sokolov DA, Yurkevich NV, Stepanova MV. Experience in the use of the fertile soil layer for the reclamation of non-ferrous metallurgy waste (Kemerovo region). Interexpo GEO-Siberia. 2022;2(2):347–354. (In Russ.). https://doi.org/10.33764/2618-981X-2022-2-2-347-354
- Kondratenko YeP, Starovoytova YeV, Starovoytov AV, Shaydulina TB, Soboleva OM. The floristic composition of segetal vegetation in spring barley crops. Bulletin of Altai State Agricultural University. 2020;185(3):35–47. (In Russ.).
- Zenova GM, Manucharova NA, Zvyagintsev DG. Extremophilic and extremotolerant actinomycetes in different soil types. Pochvovedenie. 2011;(4):457–478. (In Russ.).
- Morozkina EV, Slutskaya ES, Fedorova TV, Golubeva LI, Koroleva OV, Tugay TI. Extremophilic microorganisms: Biochemical adaptation and biotechnological application (review). Applied Biochemistry and Microbiology. 2010;46(1):5–20. (In Russ.).
- Yumatova MA. Studying of physiological and biochemical characteristics of Bacillus subtilis bacteria. Herald of Science and Education. 2019;60(6–2):5–8. (In Russ.).
- Popova АМ, Mischenko TA. Elements of bacteriology at school. Syktyvkar University Bulletin. Series 2: Biology. Geology. Chemistry. Ecology. 2020;16(4):98–108. (In Russ.).
- Belkina DD, Yurchenko EG, Karpova DV. Bacterial and yeast communities of vine of Moldova and Merlot grape varieties. Scientific works of the North Caucasian Federal Scientific Center for Horticulture, Viticulture, and Winemaking. 2021;33:69–73. (In Russ.). https://doi.org/10.30679/2587-9847-2021-33-69-73
- Pereira APM, Stradiotto GC, Freire L, Alvarenga VO, Crucello A, Morassi LLP, et al. Occurrence and enumeration of rope-producing spore forming bacteria in flour and their spoilage potential in different bread formulations. LWT. 2020;133. https://doi.org/10.1016/j.lwt.2020.110108
- Siddiqui F, Gupta A, Purwar S, Saigal S, Sharma JP. A prospective study to reduce turnaround time of microbiologically positive blood cultures in patients with sepsis in intensive care unit. Indian Journal of Medical Microbiology. 2022;40(4):541–546. https://doi.org/10.1016/j.ijmmb.2022.07.014
- Sun H, Zhang J, Liu W, E W, Wang X, Li H, et al. Identification and combinatorial engineering of indole-3-acetic acid synthetic pathways in Paenibacillus polymyxa. Biotechnology for Biofuels and Bioproducts. 2022;15. https://doi.org/10.1186/s13068-022-02181-3
- Lei H, Peng Z, Yigang H, Yang Z. Vegetation succession and soil infiltration characteristics under different aged refuse dumps at the Heidaigou opencast coal mine. Global Ecology and Conservation. 2015;4:255–263. https://doi.org/10.1016/j.gecco.2015.07.006
- Siddiqui F, Gupta A, Purwar S, Saigal S, Sharma JP. A prospective study to reduce turnaround time of microbiologically positive blood cultures in patients with sepsis in intensive care unit. Indian Journal of Medical Microbiology. 2022;40(4):541–546. https://doi.org/10.1016/j.ijmmb.2022.07.014
- Zandanova TN, Gogoleva PA. Selection of medium for obtaining bacterial concentrate of microbial consortium. Bulletin of KSAU. 2018;140(5):227–232. (In Russ.).
- Borisova EE. The role of red clover in the ecologization and biologization of agriculture. Symbol of Science: International Scientific Journal. 2016;16(4–4):56–61. (In Russ.).
- Luziatelli F, Ficca AG, Bonini P, Muleo R, Gatti L, Meneghini M, et al. A genetic and metabolomic perspective on the production of indole-3-acetic acid by Pantoea agglomerans and use of their metabolites as biostimulants in plant nurseries. Frontiers in Microbiology. 2020;11. https://doi.org/10.3389/fmicb.2020.01475
- Lee K-E, Radhakrishnan R, Kang S-M, You Y-H, Joo G-J, Lee I-J, et al. Enterococcus faecium LKE12 cell-free-extract accelerates host plant growth via gibberellin and indole-3-acetic acid secretion. Journal of Microbiology and Biotechnology. 2015;25(9):1467–1475. https://doi.org/10.4014/jmb.1502.02011
- Snak A, Vendruscolo ECG, dos Santos MF, Fiorini A, Mesa D. Genome sequencing and analysis of plant growth-promoting attributes from Leclercia adecarboxylata. Genetics of Microorganisms. 2021;44(1). https://doi.org/10.1590/1678-4685-GMB-2020-0130
- Shurigin V, Egamberdieva D, Li L, Davranov K, Panosyan H, Birkeland N-K, et al. Endophytic bacteria associated with halophyte Seidlitzia rosmarinus Ehrenb. ex Boiss. from saline soil of Uzbekistan and their plant beneficial traits. Journal of Arid Land. 2020;12(5):730–740. https://doi.org/10.1007/s40333-020-0019-4
- Jahn L, Hofmann U, Ludwig-Müller J. Indole-3-acetic acid is synthesized by the endophyte Cyanodermella asteris via a tryptophan-dependent and -independent way and mediates the interaction with a non-host plant. International Journal of Molecular Sciences. 2021;22(5). https://doi.org/10.3390/ijms22052651
