

Аннотация
Haploid technologies are used to create homozygous lines for accelerated breeding. We aimed to optimize the technology for using the isolated microspore culture in vitro to obtain doubled haploids of the carrot (Daucus carota L.).We studied two carrot varieties with different responsiveness to embryogenesis, Altajskaya lakomka and Breeding line 17. Carrot microspores were isolated from buds and cultivated in liquid nutrient media supplemented with an antibiotic and activated carbon in vitro. They were exposed to different thermal treatments.
The experiment showed the benefits of combining cold pre-treatment of buds (5°C for 1 day) with heat shock of isolated microspores in vitro (32°C for 2 days). The induction of embryogenesis on the NLN-13 medium was twice as high as on the MSm-13 medium. The use of 1% activated carbon in 0.5% agarose increased the yield of embryoids by more than 1.5 times. 100 mg/L of ampicillin was found to be the most efficient concentration. After 30 days of cultivation under optimized conditions, the yield was 161.3 and 44.0 embryoids per Petri dish for the cultivar Altajskaya lakomka and Breeding line 17, respectively.
The induction of carrot embryogenesis is determined by the type and duration of thermal stress, the composition of the nutrient medium, the use of activated carbon as a sorbent, the addition of β-lactam antibiotics, and the type of explant exposed to thermal treatment. Our technology enabled us to obtain homozygous doubled haploid lines of carrots during a year, and these lines were included in the breeding process to create F1 hybrids.
Ключевые слова
Daucus carota, culture medium, ampicillin, haploids, table carrot, penicillin, cold and heat stress, cefotaxime, embryogenesisВВЕДЕНИЕ
The carrot (Daucus carota L.) is an economically important root vegetable of the Apiaceae Lindley (Umbelliferae Juss.) family. It is rich in carotene (provitamin A) and contains an essential oil. Since the carrot is a cross-pollinated biennial crop, it takes a lot of time and effort to obtain inbred lines and select parental pairs for crossing. Moreover, it is difficult to achieve a high level of homozygosity by using traditional breeding methods.
Over the last 100 years, scientists have looked for ways to create genetic diversity by using doubled haploids. So far, they have described protocols for obtaining doubled haploids for 384 species, including the culture of isolated microspores in vitro for Apiaceae vegetables. This technology is based on the ability of microspores to switch from gametophytic to sporophytic development under conditions in vitro. Exposed to various stress factors, they form embryoids from which doubled haploids are obtained. This approach speeds up the creation of initial homozygous material (doubled haploid lines) for selection and expands the range of genetic recombinant forms, including those with recessive traits, thus facilitating the search for rare genotypes [5].
Embryogenesis induction protocols have been developed to obtain haploid and doubled haploid plants of the Apiaceae family (cumin, carrot, celery, fennel, and dill) using the cultures of unpollinated ovules, anther, and isolated microspores [6–10].
However, the practical application of unpollinated ovules and anther cultures is limited by the low efficiency of embryogenesis [11]. Moreover, there is a risk of obtaining callus and embryoids from somatic tissues during the cultivation of anthers and unpollinated ovules [12]. Therefore, isozyme or DNA analysis is required to confirm homozygosity [7]. Yet, the culture of isolated microspores excludes the interference of somatic tissues during embryogenesis.
The callus culture obtained from isolated carrot microspores was first described by Matsubara et al. in 1995, followed by studies on the regeneration of doubled haploids in the carrot [10, 13, 14]. Federal State Budgetary Scientific Institution Federal Scientific Vegetable Center (Russia) studied the process of embryogenesis and developed a method for obtaining doubled haploid plants from the culture of isolated carrot microspores [15, 16].
Many factors affect the efficiency of embryogenesis in the microspore culture in vitro, including genotype, microspore development stage, donor plant growth conditions, medium composition, and cultivation conditions [5].
Stress factors such as cold and heat treatment have a direct influence on microspores and can stimulate the induction of embryogenesis. Nitsch and Norreel first described the use of cold pre-treatment for Datura innoxia L. anthers [17]. Its positive effect was presumably associated with slowing down degradation in anther tissues. Such treatment protects microspores from toxic compounds released by degrading anthers and ensures the survival of most embryogenic microspores. Cold pre-treatment of buds has also been shown to increase the frequency of endoreduplication. This stimulates the spontaneous doubling of haploid plants and significantly enhances the efficiency of doubled haploid production. Lowering the temperature by 10°C prior to the isolation of microspores from the donor plants of the Brassica species slows down the plant’s growth and allows for a longer time when buds can be selected at a certain stage of development. Cold treatment of donor plants improves the embryogenic responsiveness of microspores. A similar phenomenon was observed in plants of the Brassica family [18].
Cold pre-treatment is mainly used for buds, while heat treatment (33–37°C) is often used to induce microspores in the culture in vitro. In anther culture, cold (4°C) and heat (32°C) pre-treatments had a positive effect on callus formation and embryogenesis of some carrot genotypes [11]. Various cold treatment schemes (1–3 days at 4°C) had a negative effect on most carrot varieties, increasing the induction of embryogenesis in only some of them [14, 15].
The composition of the nutrient medium affects embryogenesis efficiency. Such media as ½ NLN-13, NLN-13, MSm-13, ½ MS, and B5 were used to obtain embryoids from carrot microspores. Matsubara et al. supplemented ½ MS with 2,4-D (1 mg/L), BAP (1 mg/L), sucrose (100 g/L), and NLN-13 [13]. Gorecka et al. and Kiszczak et al. used the nutrient medium B5 supplemented with L-glutamine (500 mg/L), L-serine (100 mg/L), 2,4-D (0.1 mg/L), NAA (0.1 mg/L), and sucrose (100 g/L) [10, 19]. Li et al. obtained carrot plants on the NLN-13 medium containing 2,4-D (0.1 mg/L), NAA (0.1 mg/L), and activated carbon [14]1.
1 MSm – modified Murashige and Skoog medium [1], NLN – Nitsch, Lichter and Nitsch medium [2], MS – Murashige and Skoog medium [3], B5 – Gamborg medium [4], 2,4-D – 2,4-dichlorophenoxyacetic acid, BAP – 6-benzylaminopurine, NAA – 1-naphthaleneacetic acid.
High concentrations of ethylene released during microspore cultivation can have a toxic effect on the normal development of embryoids. According to literature, activated carbon is used as an adsorbing agent for ethylene [20]. In addition, it can absorb some phenols produced by damaged tissues. Shumilina et al. reported a positive effect of activated carbon on the development of somatic embryoids of the Brassica family [18].
Endogenous and exogenous microbial contamination of the culture is one of the main problems that can occur during androgenesis, which may result in the loss of valuable and rare specimens in vitro. β-lactam antibiotics are most commonly used to prevent that. They inhibit the synthesis of peptidoglycan, the main component of the bacterial cell wall, thereby exerting a bactericidal effect. Some antibiotics can have a regenerative effect, like plant growth regulators. This ability is associated with the similarity of their chemical structure with that of some phytohormones [21]. β-lactam antibiotics are similar to auxins, since their molecular structures contain four-membered (β-lactam ring) and fivemembered (pyrrole) nitrogenous heterocycles [22, 23].
The microspore culture in vitro is an advanced technology that speeds up the selection process by rapidly generating homozygous doubled haploid lines. However, it requires further research to ensure a high level of efficiency. We aimed to study the effect of various factors on the yield of carrot (D. carota L.) embryoids in the microspore culture in vitro. The factors included cold and heat treatment of explants and isolated microspores, the composition of nutrient media, as well as the presence of an antibiotic and activated carbon in the medium.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Plant material and growth conditions. Experimental samples of the carrot (Daucus carota L.) included those of the cultivar Altajskaya lakomka and Breeding line 17.
The roots were vernalized at 4°C for 4 months and then planted in a climatic chamber with 9000 lux illumination, a 16/8 h (day/night) photoperiod, and a permanent temperature of 19°C.
Umbels were collected from each sample at the beginning of flowering at an optimal stage of development according to their morphology (flat). To determine the stage of microspore development, buds from the two outer rows of umbels were placed in a mixture of 96% ethanol and glacial acetic acid (3:1 by volume) for 1–2 h and then washed with 70% ethanol. The fixed anthers were crushed in a drop of 2% acetocarmine on a glass slide and quickly heated over a burner flame (not boiled). When all the tissues were removed, they were covered with a glass slide and examined under an Axio Imager A2 light microscope (Carl Zeiss Microscopy GmbH, Germany). The stage of development was determined by the arrangement of vacuoles and nuclei. For cultivation, buds were selected with microspores mainly at the late vacuolized mononuclear stage and pollen grains at the early twocelled stage (a moderate amount) [15].
Microspore culture. Carrot buds from the two outer rows of umbels were sterilized in 96% ethanol (30 s) and then in a 50% aqueous solution of the Belizna preparation (containing 5% sodium hypochlorite) with 2–3 drops of Tween-20 (5 min). After that, they were washed three times in sterile distilled water. Microspores were isolated and cultivated according to an optimized method developed for the Brassicaceae family [24]. The microspore suspension density was determined using a Goryaev counting chamber (MiniMed, Bryansk, Russia). The suspension with a density of 2×104 was used for incubation.
The isolated microspores were cultivated in 6-cm disposable Petri dishes (filled with 3–4 drops of agarose with activated carbon) on a 5-cm3 liquid nutrient medium, NLN-13 or MSm-13 (pH = 5.8), containing 13% sucrose [1, 2]. The medium was supplemented with ampicillin (100 mg/L) and plant growth regulators, 1-naphthaleneacetic acid and 2,4-D (1 mg/L). The cultivation took place in the dark in a Binder BF 260 incubator (Tuttlingen, Germany) at a temperature according to the experimental plan.
We counted the number of embryoids (per Petri dish) formed on days 28–32 of cultivation.
Effect of thermal treatment on isolated microspores. The culture of isolated microspores in vitro on NLN-13 was exposed to cold treatment at 5°C for 1–3 days, heat treatment at 32°C for 1–2 days, and joint treatment with cold at 5°C for 2 days and heat at 32°C for 2 days [2]. The experiment included seven samples and was performed under different conditions in three repetitions:
1. 25°C throughout the entire period (control);
2. 5°C for 1 day, then 25°C;
3. 5°C for 2 days, then 25°C;
4. 5°C for 3 days, then 25°C;
5. 32°C for 1 day, then 25°C;
6. 32°C for 2 days, then 25°C; and
7. 5°C for 2 days, 32°C for 2 days, then 25°C.
Effect of thermal treatment on explants and isolated microspores. Two types of explants were obtained from four umbels of each carrot sample – buds separated from inflorescences and whole inflorescences. They were pre-treated with cold at 5°C for 1– 2 days. Then, the culture of isolated microspores in vitro on NLN-13 was exposed to heat shock at 32°C for 2 days [2].
The experiment was performed in seven repetitions and three replicates, namely:
1. 32°C for 2 days, then 25°C (control);
2. 5°C for 1 day, 32°C for 2 days, then 25°C; and
3. 5°C for 2 days, 32°C for 2 days, then 25°C.
Effect of the nutrient medium composition. Two liquid nutrient media were used for the experiments, NLN-13 and MSm-13 (pH = 5.8), containing 13% sucrose and supplemented with ampicillin (100 mg/L) and plant growth regulators NAA and 2.4-D (1 mg/L) [1, 2].
The experiment was performed in seven repetitions.
Effect of activated carbon. 0.5 g of agarose and 1 g of activated carbon were dissolved in 100 cm3 of distilled water. The preparation was autoclaved and dropped into Petri dishes, 3–4 drops each. Carrot buds were pre-treated with cold at 5°C for 1 day. Then the culture of isolated microspores in vitro on NLN-13 was subjected to heat treatment at 32°C (2 days) and then at 25°C [2].
The experiment was performed in two biological replicates: without activated carbon (control) and with activated carbon.
Three analytical replicates were performed for each experimental variant.
Effect of antibiotics. Ampicillin (Biosintez, Russia), cefotaxime (Sintez, Russia), and penicillin (Biosintez, Russia) were used in a lyophilized injection form in sterile 1-g vials. To prepare an antibiotic solution, sterile water (10 cm3) was added to a sterile antibiotic (1 g). Then, the solution was aliquoted by 1 cm3 and stored at –20°C. Carrot buds were pre-treated with cold at 5°C for 1 day. The culture of isolated microspores in vitro on NLN-13 was subjected to heat treatment at 32°C (2 days) followed by 25°C [2]. The antibiotics were added to the nutrient medium according to the experimental plan.
The phytotoxicity of antibiotics was determined in four replicates, namely: without antibiotic (control), cefotaxime (100 mg/L), ampicillin (100 mg/L), and penicillin (100 mg/L).
The optimal concentration of ampicillin was determined in five replicates, namely: without antibiotic (control), ampicillin (50 mg/L), ampicillin (100 mg/L), ampicillin (200 mg/L), and ampicillin (1000 mg/L).
Three analytical replicates were performed for each experimental variant.
All the reagents, except for the antibiotics, were produced by Sigma-ALDRICH (USA) and were labeled as “suitable for the plant cell culture”.
Statistical data analysis. Experimental data were processed in Microsoft Excel 2010. One-way and twoway analysis of variance (ANOVA) was performed on the basis of the Fisher test. The least significant difference (LSD) was evaluated using Student’s t-test with a probability of 0.95. Experimental data are given as means ± error of the mean.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Effect of cold and heat treatment of isolated carrot microspores and explants on the induction of embryogenesis in culture in vitro. The process of primary and secondary embryogenesis in the microspore culture of the carrot (Daucus carota L.) was described earlier by Shmykova et al. [15]. The technology for obtaining doubled haploids in the microspore culture in vitro has been developed for individual carrot genotypes [5, 14]. However, due to a large number of factors that influence embryogenesis and regeneration, these processes need to be continuously reviewed, taking into account innovations and genotypes.
In 2020–2021, we conducted an experiment in two stages to assess carrot microspore responsiveness to embryogenesis at different temperatures. At the first stage (2020), isolated microspores in the culture in vitro were subjected to heat treatment (Table 1). At the second stage (2021), explants were pre-treated with cold and then isolated microspores in the culture in vitro were exposed to heat (Table 2).
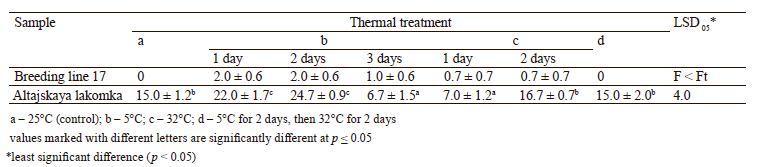

At the first stage of the experiment in 2020, we studied the effect of cold and heat treatments, as well as their combinations, on the embryogenic induction of microspores by the carrot samples (Table 1). We found that cold treatment at 5°C stimulated the formation of carrot embryoids. The best results were obtained by cold treatment during 1 and 2 days (22.0 and 24.7 embryoids per Petri dish, respectively). However, longer cold treatment (up to 3 days) reduced the induction of embryogenesis to 6.7 embryoids per Petri dish, which was half the control number of 15.0 embryoids per Petri dish.
The heat stress at 32°C did not have a positive effect on the induction of embryogenesis in the microspore culture in vitro. In particular, 1 and 2 days of heat treatment produced only 7.0 and 16.7 embryoids per Petri dish, respectively, which corresponded to the control level. A combination of cold (5°C) and heat (32°C) treatments during 2 days had no significant effect either, with the number of embryoids reaching the control level (15.0 per Petri dish). According to Table 1, the Breeding line 17 samples showed little responsiveness to the experimental conditions (0–2.0 embryoids per Petri dish) and produced results with no statistically significant differences (F < Ft). The Altajskaya lakomka samples were much more responsive, with 6.7– 24.7 embryoids per Petri dish. In particular, we found this cultivar responsive to cold treatment at 5°C for 1– 2 days. Therefore, this treatment regime was used at the next stage of the experiment the following year.
At the second stage of the experiment in 2021, we combined cold pre-treatment of explants (5°C for 1 and 2 days) with heat treatment of isolated microspores (32°C for 2 days). This combination increased embryogenic induction to 28.9 and 58.2 embryoids per Petri dish for the Breeding line 17 and cultivars Altajskaya lakomka, respectively. A longer cold pre-treatment of explants (2 days) resulted in the embryogenic induction at the control level (Table 2).
The cultivar Altajskaya lakomka showed high responsiveness to the experimental conditions at the second stage, while the Breeding line 17 samples formed embryoids only after a 1-day cold pre-treatment of explants. Yet, even with cold pre-treatment, Breeding line 17’s embryogenic activity was twice as low as that of the Altajskaya lakomka (28.9 and 58.2 embryoids per Petri dish, respectively). Moreover, a 2-day cold pretreatment of Breeding line 17 explants at 5°C was totally ineffective, producing 0 embryoids per Petri dish.
Cold stress can stimulate embryogenic induction. In our experiments, however, its positive effect at 5°C was short-lived. In particular, only one day of cold treatment increased the yield of embryoids, with longer treatments leading to decreased embryogenic induction. This is probably associated with the fact that microspores quickly reached a stage of development optimal for embryogenesis. Our results were consistent with those from studies on carrot anthers [11, 14].
Responsiveness to microspore embryogenesis greatly depends on the genotype of a donor plant. Differences in embryogenic induction are also determined by the year of the study [14]. In our experiment, the cultivar Altajskaya lakomka proved highly responsive to embryogenesis, while Breeding line 17 revealed low embryogenic potential. We believe it was due to the fact that Breeding line 17 microspores were at an unsuitable stage of development at the time of the experiment. It may also be that the 24-h duration of the cold stress (5°C) was too long for the carrot buds and it needs to be shortened.
In addition, the genotype of a donor plant affected embryogenic induction. In particular, a combination of cold treatment of the Altajskaya lakomka buds at 5°C for 2 days and the subsequent heat treatment at 32°C for 2 days produced different numbers of embryoids in 2020 and 2021 (15.0 and 42.2 per Petri dish, respectively) (Tables 1 and 2). This could be because the experiments were carried out during mass flowering and inflorescences were collected from different donor plants.
Effect of the nutrient medium composition. We found that embryogenetic induction differed depending on the composition of the nutrient medium. Both carrot varieties, Breeding line 17 and Altajskaya lakomka (Table 3), showed a twice higher induction potential on the NLN-13 medium (13.8 and 64.6 embryoids per Petri dish, respectively) than on the MSm-13 medium (5.8 and 28.2 embryoids per Petri dish, respectively).
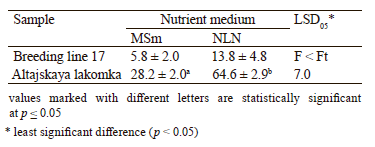
The differences in the composition of the nutrient medium for Breeding line 17 samples were statistically insignificant.
The choice of the nutrient medium has a significant effect on the efficiency of embryogenesis. Matsubara et al. were the first to observe the division of carrot microspores on a liquid nutrient medium ½ MS supplemented with 2,4-D (1 mg/L) and BAP (1 mg/L) [3, 13]. However, no embryoids were obtained in that study. Gorecka et al. successfully used the B5 medium supplemented with glutamine (500 mg/L), L-serine (100 mg/L), 2,4-D (0.1 mg/L), and NAA (0.1 mg/L) for the microspore culture [10]. Li et al. induced carrot microspores on the NLN-13 medium supplemented with 2,4-D (0.1 mg/L) and NAA (0.1 mg/L) [2, 4, 14]. Vjurtts et al. reported a successful formation of carrot embryoids on the ½ NLN-13 and MS-13 media with various additives [2, 3, 16]. In later studies, Shmykova et al. observed the induction of embryogenesis in carrots on the ½ NLN-13 medium supplemented with 2,4-D (0.2 mg/L) and kinetin (0.2 mg/L) [2, 15].
In our study, the NLN-13 medium yielded twice as many embryoids per Petri dish as the MSm-13 medium. This pattern was observed for both varieties. In earlier experiments conducted on other carrot genotypes, the NLN-13 medium also showed the highest induction of embryogenesis [16]. These results are due to the fact that NLN-13 was developed directly for cultivating rapeseed microspores, with stress effects taken into account. For this, the medium contained reduced glutathione and an increased content of boron ions, which triggered cell division. The MSm-13 medium, however, was used to induce gynogenesis and obtain carrot embryoids from callus cells [1, 12]. Therefore, it is better suited for further cultivation of the embryoids obtained on NLN-13 and for growing seedlings of carrot plants [2].
Effect of the explant type. We used two types of explants: buds separated from inflorescences and whole inflorescences cut and placed in a glass of water (Table 4). The explants were pre-treated with cold, after which microspores were isolated and introduced into the culture in vitro.
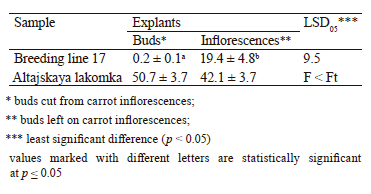
The effect of cold pretreatment on the induction of embryogenesis depended on the carrot genotype. Particularly, the Altajskaya lakomka samples showed the best results when microspores were isolated from carrot buds (50.7 embryoids per Petri dish). Yet, the influence of the explant type was insignificant for this cultivar. As for the Breeding line 17 samples, the largest number of embryoids (19.4 per Petri dish) was produced by the microspores isolated from inflorescences. The differences between the two types of explants were significant for this cultivar.
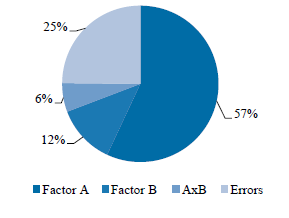
According to the two-way analysis of variance with repetitions, the composition of the nutrient medium had the biggest effect on the efficiency of carrot embryogenesis in the microspore culture in vitro, accounting for 57% (Fig. 1). The effect of thermal treatment amounted to 12%, while the combined effect of these two factors was only 6%.
The influence of these factors on the efficiency of embryoid formation was not unambiguous, which may be due to differences in the carrot genotypes. However, the one-way analysis of variance revealed no significant differences. A larger number of samples and factors to be studied might give a better understanding of how different types of explants affect the induction of embryogenesis.
Effect of activated carbon. Adding activated carbon powder to a liquid nutrient medium causes carrot microspores to stick together and impairs their further development (Fig. 2). Therefore, we used a preparation of activated carbon with agarose.
The use of activated carbon as an adsorbing agent in the lower layer of the liquid nutrient medium increased the formation of embryoids in the microspore culture in vitro 1.5–1.8 times, compared to the carbon-free control (Table 5, Fig. 3). The cultivar Altajskaya lakomka proved more responsive and yielded 4–5 times more embryoids that Breeding line 17.

The one-way analysis of variance showed that the addition of activated carbon to the nutrient medium was a significant factor, the data belonging to different groups.
Earlier studies reported that adding activated carbon to the bottom layer of a two-layer nutrient medium increased the rate of embryoid formation in Brassica napus. Gland et al. suggested that activated carbon removes toxic substances released by inactive microspores, stimulating embryoids to develop [25]. In addition, it may adsorb phenolic compounds that are released from damaged tissues during cultivation. In a study on embryogenesis and regeneration of wheat seedlings (Triticum aestivum L.), activated carbon increased the percentage of morphogenic callus and the number of green meristematic foci. The largest number of green shoots was obtained on the medium containing activated carbon (0.5 g/L) [26].
We found that supplementing the nutrient medium with a 1% suspension of activated carbon in 0.5% agarose intensified embryogenesis in the carrot microspore culture in vitro. Guo and Pulli showed a positive effect of adding activated carbon in agarose [27]. In a study by Prem et al., activated carbon was added without agarose and had a negative effect [28]. The authors suggested that carbon particles stuck to the microspores and embryoids, thereby preventing their growth and development, which was confirmed by our experiments.
According to our results, using activated carbon in the microspore culture of more responsive genotypes significantly increases the formation of embryoids. Even the less responsive Breeding line 17 showed a slight increase in embryoids compared to the control (without activated carbon).
Effect of β-lactam antibiotics. When using the standard protocol for the cultivation of microspores, we found a serious loss of starting material in some samples due to endogenous microbial contamination in vitro. In the experiments without antibiotics, microbial contamination and a total loss of starting material in some samples were observed on the third day. Yet, adding ampicillin to the nutrient medium, even at the lowest concentration of 50 mg/L, prevented microbial contamination (Fig. 4).
The phytotoxicity of various antibiotics (cefotaxime, ampicillin, penicillin) was determined in the microspore culture in vitro of the highly responsive cultivar Altajskaya lakomka according to previous studies on cabbage crops [23]. The sample without antibiotics (control) had bacterial contamination (no embrioids), which led to complete inhibition of the microspore culture. Ampicillin provided the largest number of carrot embryoids (161.3 per Petri dish), while cefotaxime reduced this number 7 times (23.3 embryoids per Petri dish). The addition of penicillin suppressed the division of microspores and the formation of embryoids (Fig. 5).

To study the induction effect of β-lactam antibiotics on the embryoid yield, we carried out an experiment with various concentrations of ampicillin as the least phytotoxic antibiotic. We found the largest number of carrot embryoids (130.5 per Petri dish) on the nutrient medium with 100 mg/L of ampicillin (Table 6). The minimum concentration of 50 mg/L did not stimulate the embryogenesis significantly (16.0 embryoids per Petri dish). Yet, concentrations exceeding 100 mg/L suppressed the development of carrot embryoids (11.8 per Petri dish).

The β-lactam antibiotics proved effective in the culture of isolated carrot microspores in vitro. Except for their high antibacterial activity, they should not have a negative effect on the induction and development of embryoids. Earlier studies showed that antibiotics can induce regeneration, but at high concentrations they inhibit the development of explants due to their toxicity [22, 23, 29].
Grzebelus and Skop, who studied the effect of β-lactam antibiotics on regeneration in the carrot protoplast culture, found that cefotaxime had a stimulating effect at a concentration of 400–500 mg/L [21]. According to Meng et al., ampicillin and cefotaxime had a selective effect on the frequency of regeneration of Chinese cabbage somatic embryoids [22]. In particular, cefotaxime inhibited re-differentiation of shoots and roots, while ampicillin stimulated their differentiation. In a study by Gerszberg and Grzegorczyk-Karolak, ampicillin at concentrations of 100–400 mg/L not only had an antimicrobial effect, but also regenerated tomato buds, while cefotaxime at a concentration of over 200 mg/L had a negative effect [29].
Despite the widespread application of antibiotics in various methods, their use in the microspore culture in vitro is an innovation. Therefore, it is not included in the standard protocol for producing doubled haploid plants by androgenesis. Yet, we have found a few studies on wheat, triticale, and rapeseed. Asif et al. found that cefotaxime at a concentration of 100 mg/L had a stimulating effect on the embryogenesis of wheat and triticale in the microspore culture in vitro [30]. Other authors reported its inducing effect on the embryogenesis and regeneration of rapeseed at certain concentrations and treatment times [31].
According to our results, ampicillin at a concentration of 100 mg/L was the most effective of all the β-lactam antibiotics under study in inducing carrot embryogenesis in the microspore culture in vitro. It not only controlled unwanted microbial infection, but also significantly increased embryoid yield. Similar results were obtained earlier for cabbage crops [24].
ВЫВОДЫ
The induction of embryogenesis in the culture of isolated microspores in vitro is determined by many factors. Although there is a protocol for obtaining haploid and doubled haploid carrot plants, this process is highly dependent on the sample and microspore cultivation conditions. Therefore, this technology needs to be improved for specific genotypes.
In our study, the following factors determined the efficiency of carrot embryogenic induction: a responsive donor plant genotype (Altajskaya lakomka), cold treatment of plant buds at 5°C for 1 day and heat treatment of isolated microspores at 32°C for 2 days, the NLN-13 nutrient medium with activated carbon as an adsorbent, and ampicillin at a concentration of 100 mg/L. We optimized the technology to increase the induction of embryogenesis not only in the highly responsive sample Altajskaya lakomka, but also Breeding line 17 with low responsiveness (157.3 and 44.0 embryoids per Petri dish, respectively, after 30 days of cultivation) (Fig. 6).
Вклад авторов
Concept development: E.A. Domblides and O.V. Romanova; methodology: E.A. Domblides; software: V.A. Akhramenko; data validation: E.A. Domblides, O.V. Romanova, and A.I. Mineikina; formal analysis: E.A. Domblides, O.V. Romanova, and A.I. Mineikina; research: T.S. Vjurtts, A.I. Mineykina, O.V. Romanova, Ya.P. Tukuser, and Yu.V. Kulakov; resources: A.V. Soldatenko; data curation: T.S. Vjurtts, A.I. Mineykina, and O.V. Romanova; the initial draft: E.A. Domblides, O.V. Romanova, and A.I. Mineykina; reviewing and editing: E.A. Domblides, O.V. Romanova, and A.I. Mineykina; visualization: Yu.V. Kulakov; project management and control: A.V. Soldatenko.ФИНАНСИРОВАНИЕ
The study was funded by the Ministry of Science and Higher Education of the Russian Federation (Minobrnauka) within State Assignment No. FGGF-2019-0001 “Developing a technology for accelerated breeding of agricultural plants based on reproductive biotechnology. Experimental haploidy to accelerate the breeding schemes for vegetable crops”.СПИСОК ЛИТЕРАТУРЫ
- Masuda K, Kikuta Y, Okazava Y. A revision of the medium for somatic embryogenesis in carrot suspension culture. Journal of the Faculty of Agriculture, Hokkaido University. 1981;60(3):183–193.
- Lichter R. Induction of haploid plants from isolated pollen of Brassica napus. Zeitschrift für Pflanzenphysiologie. 1982;105(5):427–432. https://doi.org/10.1016/S0044-328X(82)80040-8
- Murashige T, Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiologia Plantarum. 1962;15(3):473–497. https://doi.org/10.1111/j.1399-3054.1962.tb08052.x
- Gamborg OL, Miller RA, Ojima K. Nutrients requirements of suspension cultures of soybean root cells. Experimental Cell Research. 1968;50(1):151–158. https://doi.org/10.1016/0014-4827(68)90403-5
- Segui-Simarro JM, Moreno JB, Fernández MG, Mir R. Species with haploid or doubled haploid protocols. In: Segui-Simarro JM, editor. Doubled haploid technology: Volume 1: General topics, Alliaceae, cereals. New York: Humana Press; 2021. pp. 41–103. https://doi.org/10.1007/978-1-0716-1315-3_3
- Smýkalová I, Horáček J. Caraway (Carum carvi L.): Anther culture and production of DH plants caraway. In: Segui-Simarro JM, editor. Doubled haploid technology: Volume 2: Hot topics, Apiaceae, Brassicaceae, Solanaceae. New York: Humana Press; 2021. pp. 91–102. https://doi.org/10.1007/978-1-0716-1335-1_5
- Kiszczak W, Burian M, Kowalska U, Górecka K. Production of homozygous carrot (Daucus carota L.) plants by anther culture. In: Segui-Simarro JM, editor. Doubled haploid technology: Volume 2: Hot topics, Apiaceae, Brassicaceae, Solanaceae. New York: Humana Press; 2021. pp. 113–126. https://doi.org/10.1007/978-1-0716-1335-1_7
- Dohya N, Matsubara S, Murakami K. Callus formation and regeneration of adventitious embryos from celery microscopes by anther and isolated microscope cultures. Journal of the Japanese Society for Horticultural Science. 1997;65(4):747–752. https://doi.org/10.2503/jjshs.65.747
- Ferrie AMR. Doubled haploidy for fennel (Foeniculum vulgare Mill.) and dill (Anethum graveolens L.). In: Segui-Simarro JM, editor. Doubled haploid technology: Volume 2: Hot topics, Apiaceae, Brassicaceae, Solanaceae. New York: Humana Press; 2021. pp. 103–111. https://doi.org/10.1007/978-1-0716-1335-1_6
- Górecka K, Kowalska U, Krzyzanowska D, Kiszczak W. Obtaining carrot (Daucus carota L.) plants in isolated microspore cultures. Journal of Applied Genetics. 2010;51(2):141–147. https://doi.org/10.1007/BF03195722
- Zhuang FY, Pei HX, Ou CG, Hu H, Zhao ZW, Li JR. Induction of microspores-derived embryos and calli from anther culture in carrot. Acta Horticulturae Sinica. 2010;37(010):1613–1620.
- Tyukavin GB, Shmykova NA, Monahova MA. Cytological study of embryogenesis in cultured carrot anthers. Russian Journal of Plant Physiology. 1999;46(6):767–773.
- Matsubara S, Dohya N, Murakami K. Callus formation and regeneration of adventitious embryos from carrot, fennel and mitsuba microspores by anther and isolated microspore cultures. Acta Horticulturae. 1995;392:129–137. https://doi.org/10.17660/ActaHortic.1995.392.15
- Li J-R, Zhuang F-Y, Ou C-G, Hu H, Zhao Z-W, Mao J-H. Microspore embryogenesis and production of haploid and doubled haploid plants in carrot (Daucus carota L.). Plant Cell, Tissue and Organ Culture. 2013;112(3):275–287. https://doi.org/10.1007/s11240-012-0235-5
- Shmykova N, Domblides E, Vjurtts T, Domblides A. Haploid embryogenesis in isolated microspore culture of carrots (Daucus carota L.). Life. 2021;11(1). https://doi.org/10.3390/life11010020
- Vjurtts TS, Domblides EA, Shmykova NA, Fedorova MI, Kan LYu, Domblides AS. Production of DH-plants in culture of isolated microspore in carrot. Vegetable Crops of Russia. 2017;38(5):25–30. (In Russ.). https://doi.org/10.18619/2072-9146-2017-5-25-30
- Nitsch C, Norreel B. Factors favoring the formation of androgenetic embryos in anther culture. In: Srb AM, editor. Genes, enzymes, and populations. New York: Springer; 1973. pp. 129–144. https://doi.org/10.1007/978-1-4684-2880-3_10
- Shumilina D, Kornyukhin D, Domblides E, Soldatenko A, Artemyeva A. Effects of genotype and culture conditions on microspore embryogenesis and plant regeneration in Brassica rapa ssp. Rapa L. Plants. 2020;9(2). https://doi.org/10.3390/plants9020278
- Kiszczak W, Kowalska U, Kapuścińska A, Burian M, Górecka K. Comparison of methods for obtaining doubled haploids of carrot. Acta Societatis Botanicorum Poloniae. 2017;86(2). https://doi.org/10.5586/asbp.3547
- Thomas TD. The role of activated charcoal in plant tissue culture. Biotechnology Advances. 2008;26(6):618–631. https://doi.org/10.1016/j.biotechadv.2008.08.003
- Grzebelus E, Skop L. Effect of β-lactam antibiotics on plant regeneration in carrot protoplast cultures. In Vitro Cellular and Developmental Biology – Plant. 2014;50(5):568–575. https://doi.org/10.1007/s11627-014-9626-0
- Meng Q, Liu Z, Zhang Y, Liu C, Ren F, Feng H. Effects of antibiotics on in vitro-cultured cotyledons. In Vitro Cellular and Developmental Biology – Plant. 2014;50(4):436–441. https://doi.org/10.1007/s11627-014-9595-3
- Mineykina A, Shumilina D, Bondareva L, Soldatenko A, Domblides E. Effect of beta-lactam antibiotics on microspore embryogenesis in Brassica species. Plants. 2020;9(4). https://doi.org/10.3390/plants9040489
- Domblides EA, Shmykova NA, Shumilina DV, Zayachkovskaya TV, Mineykina AI, Kozarʹ EV, et al. A technology for obtaining doubled haploids in microspore cultures of the Brassicaceae family (guidelines). Moscow: VNIISSOK; 2016. 40 p. (In Russ.).
- Gland A, Lichter R, Schweiger H-G. Genetic and exogenous factors affecting embryogenesis in isolated microspore cultures of Brassica napus L. Journal of Plant Physiology. 1988;132(5):613–617. https://doi.org/10.1016/S0176-1617(88)80264-5
- Abdollah HA, Said AGE, Khalafalla MM. Embryogenesis and plantlet regeneration optimization of wheat (Triticum aestivum L.). International Journal of Agricultural Technology. 2014;10(3):679–693.
- Guo Y-D, Pulli S. High-frequency embryogenesis in Brassica campestris microspore culture. Plant Cell, Tissue and Organ Culture. 1996;46(3):219–225. https://doi.org/10.1007/BF02307098
- Prem D, Gupta K, Agnihotri A. Effect of various exogenous and endogenous factors on microspore embryogenesis in Indian mustard (Brassica juncea (L.) Czern and Coss). In Vitro Cellular and Developmental Biology – Plant. 2005;41(3):266–273. https://doi.org/10.1079/IVP2005636
- Gerszberg A, Grzegorczyk-Karolak I. Influence of selected antibiotics on the tomato regeneration in in vitro cultures. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2019;47:558–564. https://doi.org/10.15835/nbha47311401
- Asif M, Eudes F, Randhawa H, Amundsen E, Yanke J, Spaner D. Cefotaxime prevents microbial contamination and improves microspore embryogenesis in wheat and triticale. Plant Cell Reports. 2013;32(10):1637–1646. https://doi.org/10.1007/s00299-013-1476-4
- Ahmadi B, Shariatpanahi ME, Ojaghkandi MA, Heydari AA. Improved microspore embryogenesis induction and plantlet regeneration using putrescine, cefotaxime and vancomycin in Brassica napus L. Plant Cell, Tissue and Organ Culture. 2014;118(3):497–505. https://doi.org/10.1007/s11240-014-0501-9
