

Аннотация
Medicinal plants are sources of natural antioxidants. Acting as reducing agents, these substances protect the human body against oxidative stress and slow down the aging process. We aimed to study the effects of bioactive substances isolated from medicinal plants on the lifespan of Caenorhabditis elegans L. used as a model organism.High-performance liquid chromatography was applied to isolate bioactive substances from the extracts of callus, suspension, and root cultures of meadowsweet (Filipendula ulmaria L.), ginkgo (Ginkgo biloba L.), Baikal skullcap (Scutellaria baicalensis L.), red clover (Trifolium pretense L.), alfalfa (Medicágo sativa L.), and thyme (Thymus vulgaris L.). Their effect on the lifespan of C. elegans nematodes was determined by counting live nematodes treated with their concentrations of 10, 50, 100, and 200 µmol/L after 61 days of the experiment. The results were recorded using IR spectrometry.
The isolated bioactive substances were at least 95% pure. We found that the studied concentrations of trans-cinnamic acid, baicalin, rutin, ursolic acid, and magniferin did not significantly increase the lifespan of the nematodes. Naringenin increased their lifespan by an average of 27.3% during days 8–26. Chlorogenic acid at a concentration of 100 µmol/L increased the lifespan of C. elegans by 27.7%. Ginkgo-based kaempferol and quercetin, as well as red clover-based biochanin A at the concentrations of 200, 10, and 100 µmol/L, respectively, increased the lifespan of the nematodes by 30.6, 41.9, and 45.2%, respectively.
The bioactive substances produced from callus, root, and suspension cultures of the above medicinal plants had a positive effect on the lifespan of C. elegans nematodes. This confirms their geroprotective properties and allows them to be used as anti-aging agents.
Ключевые слова
Plants, antioxidants, callus culture, suspension culture, root culture, nematodes, agingВВЕДЕНИЕ
According to the WHO, average life expectancy is steadily increasing worldwide [1]. Over the last 20 years, it has grown by 6 years as a result of advances in science and medicine. However, behind these advances is an increase in diseases associated with aging, which has become a serious problem of public health in the 21st century. Aging is a process that affects the entire human body, in particular its cardiovascular, nervous, digestive, and immune systems.
The aging process is directly related to oxidative stress. Age-related diseases cause structural changes in mitochondria, as well as changes in the functions of the electron transport chain, which ultimately leads to oxidative stress. The cardiovascular system is particularly susceptible to this effect, which explains the increase in cardiovascular diseases in the elderly [2–5].
The problem of aging is widely covered by the free-radical theory of aging developed by Denham Harman in the 1950s [6]. According to this theory, the body’s defense mechanisms stop responding to damage caused by reactive oxygen species, resulting in the deterioration of cellular homeostasis, energy imbalance, and mitochondrial insufficiency [7].
The molecules of reactive oxygen species include nitric oxide, hydrogen peroxide, monoxide radicals, superoxide anions, tocopherols, ascorbic acid, and polyphenols [8]. Reactive oxygen species alter cellular activities such as cell survival, stress responses, and inflammation. They are also involved in muscle contractions, regulate vascular tone, as well as determine bactericidal and bacteriostatic activity [9]. However, their increase leads to oxidative stress, disrupting the balance of antioxidants and prooxidants [10]. This causes damage to macromolecules (lipids, DNA, and proteins) and subsequently to whole cells, tissues, and organs [4]. Higher concentrations of reactive oxygen species in the body promote inflammation, which, in turn, can accelerate the formation of their molecules [11]. Therefore, it is very important to maintain a balance between antioxidants and pro-oxidants.
Antioxidants can react with free radicals and neutralize them by causing them to decay. The human body has three lines of defense against oxidative stress. The first line consists of body enzymes such as superoxide dismutase, catalase, glutathione peroxidase, glutathione S-transferase, and glutathione reductase [12]. They prevent cell damage by scavenging free radicals that cause premature aging and age-related disorders. The first line also includes non-enzymatic molecules in the blood plasma such as transferrin, ferritin, ceruloplasmin, and albumin [13]. These preventative antioxidants inhibit the formation of new reactive oxygen species by binding transition metal ions (for example, copper and iron). The second line of defense is represented by non-enzymatic antioxidants that provide intermediate protection against oxidative radicals. The third line of defense serves to regenerate biomolecules damaged by oxidative stress [14].
However, protection against oxidative stress should not be limited to the protective function of the biological system itself. Noteworthily, one antioxidant molecule is capable of reacting with only one oxidizing radical. Therefore, there is a need to replenish antioxidant molecules, including the use of supplements. There are three groups of exogenous antioxidants: mineral elements, nutritional antioxidants (carotenoids, vitamins E and C), and natural antioxidants derived from natural sources commonly known as phytochemicals or phytonutrients [15].
Synthetic antioxidants have been widely used until recently, but there is some doubt as to their usefulness and safety. According to some studies, synthetic antioxidants are ineffective against oxidative stress. Moreover, their long-term use can lead to diseases such as skin allergies, gastrointestinal and cardiovascular diseases, and even increase the risk of cancer [16]. Therefore, there is a need for thorough research into the safety of synthetic antioxidants.
The main sources of exogenous antioxidants are fruits, vegetables, cereals, etc. However, modern research is focused on traditional medicinal plants as a source of natural antioxidants [17, 18].
Since ancient times, plants have been a source of many useful substances, including exogenous antioxidants. These substances act as reducing agents that scavenge free radicals, protect the body against oxidative stress, and, as a result, maintain a balance between oxidants and antioxidants [19]. This is achieved due to the presence of polyphenols, tocopherols, carotenoids, ascorbic acid, and macromolecules (including polysaccharides and peptides), as well as essential oils [20].
Polyphenols are substances that contain a multiple number of structural units of phenol [21]. Their number affects the chemical, biological, and physical properties of polyphenolic compounds. Polyphenols are represented by flavonoids, phenolic acids, and nonflavonoids [22, 23]. Depending on the chemical structure, flavonoids are classified into flavonols, flavanones, isoflavones, anthocyanins, and flavan-3-ols. They are the most abundant class of polyphenols, with about 8000 compounds identified to date [24]. A couple of decades ago, researchers significantly increased their interest in polyphenols due to their beneficial properties for humans [25]. In particular, polyphenols curb oxidative stress and related conditions through their reductive ability to protect cellular components from oxidative damage caused by free radicals [26].
Model organisms have become an indispensable part of biological studies that would be impossible to conduct on humans for ethical or economic reasons [27]. Studies of aging and age-related diseases require an organism with a relatively short lifespan and clearly identified genetic factors to ensure reproducibility and reliability [28]. Nematodes (Caenorhabditis elegans L.), drosophila (Drosophila melanogaster L.), and yeasts (Saccharomyces cerevisiae L.) are used to test the effect of bioactive substances on lifespan [29, 30].
C. elegans is a non-parasitic, free-living nematode that feeds on various bacteria, primarily Escherichia coli [31]. This simple multicellular organism up to 1 mm long is a hermaphrodite capable of self-fertilization [32]. Its hermaphroditic structure contributes to low genetic variability [33]. C. elegans has a short life cycle (2– 4 days) and remains viable for 20–25 days at 20°C as an adult [34]. The nematode can be stored in liquid nitrogen for an almost unlimited amount of time [35].
C. elegans was first used as a model organism for biological research by Sydney Brenner in the 1965s [36]. Since then, it has been widely used to study aging and associated oxidative stress, as well as neurodegeneration and inflammation processes [37].
C. elegans has a number of advantages that explain its common uses as a model organism. Firstly, the nematode is easily cultivated and has a transparent body, which makes it easy to track the changes microscopically [33, 38–40]. Secondly, C. elegans has four organ systems that are the same as in vertebrates (nervous, digestive, immune, and reproductive), which allows for reliable and valuable conclusions [41]. Thirdly, its short lifespan (20–25 days) enables scientists to conduct rapid experiments aimed to study the effect of various substances on the lifespan [42]. Finally, the nematode’s genome is completely deciphered and easily modified, which facilitates studies of the aging process [43–46].
Plants of the Siberian Federal Okrug (Russia) are potential sources of geroprotectors – substances that can slow down the aging process [47].
Meadowsweet (Filipendula ulmaria L.) is a herbaceous perennial plant common in Russia and many European countries [48]. Extracts of this plant have anticancerous, antioxidant, and anti-inflammatory activity [49]. This activity is associated with tannins, phenolic compounds, phenolcarboxylic acids, catechins, flavonoids, essential oils, and other bioactive substances contained in the roots and flowers of the plant [50]. Many previous studies confirm its medicinal properties [51, 52]. Rutin, one of its phytochemicals, belongs to the class of natural flavonoids and is known as quercetin-3-O-rutinoside or vitamin P [53]. It has antitumorous, anticarcinogenic, and antimicrobial properties [54, 55].
Ginkgo (Ginkgo biloba L.) is a special medicinal plant that contains a variety of compounds with a unique structure due to its phylogenetic divergence from other plants. Its extract obtained by drying the leaves is used to treat many neurodegenerative diseases (memory impairment, dementia, Alzheimer’s disease) [56]. It is also widely applied as an antiinflammatory, cardioprotective, and antioxidant agent [57]. The plant’s active components are flavonoids, terpenoids, polyphenols, and organic acids [58]. Quercetin and kaempferol are two of these beneficial compounds. Quercetin is a flavonoid that has a positive effect on cardiovascular diseases, breast cancer, and ischemia [59–62]. Kaempferol is a valuable component with anticancerous, antitumorous, and antiinflammatory properties [62].
Baikal skullcap (Scutellaria baicalensis L.) belongs to the genus Lamiaceae and is still widely used in Chinese traditional medicine [63]. This plant grows in China, Russia, Mongolia, Japan, and North Korea. To date, scientists have identified 126 low-molecular weight compounds in it, mostly in its root. These compounds include flavonoids, flavonoid glycosides, and phenylethanoid glycosides [64]. The most widespread and studied of them is baicalin, which is used for various medical purposes [66].
Red clover (Trifolium pretense L.) is one of the most important representatives of the Leguminosae family, numbering over 240 species [66]. It has numerous medicinal properties and therapeutic effects on respiratory diseases, bacterial and fungal infections, tumors, and diabetes [67]. The plant is rich in isoflavones (biochanin A, genistein, trifoside), flavonoids (quercetin, kaempferol), as well as cinnamic, caffeic, and chlorogenic acids [68].
Alfalfa (Medicágo sativa L.) is a flowering plant in the Fabaceae family, which is the largest and most widespread family in the world. The genus Medicágo includes 83 species rich in alkaloids, flavonoids, naphthoquinones, and saponins [69]. Naringenin, one of its components, is a water-soluble flavonoid of great value due to its anticancerous, antioxidant, and antiinflammatory effects [70, 71].
Thyme (Thymus vulgaris L.) is an aromatic perennial flowering plant belonging to the Lamiaceae family. Its therapeutic properties are mainly associated with its essential oil that has antitussive, expectorant, antiseptic, antimicrobial, and anthelmintic effects [72]. The plant is traditionally used to treat oral, gastrointestinal, and urinary tract infections, as well as respiratory diseases (cough, bronchitis, asthma) [73]. Ursolic acid, one of its bioactive substances, is a promising agent against cancer, cardiovascular disease, brain and liver diseases, obesity, and diabetes [74, 75].
We aimed to study the effect of individual bioactive substances on the lifespan of the model organism C. elegans.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
We used individual bioactive substances isolated from the extracts of suspension, callus, and root cultures of Siberian medicinal plants. The extraction parameters are presented in Table 1.

High-performance liquid chromatography (HPLC)
was applied (Shimadzu LC-20 Prominence liquid
chromatograph, Japan) to isolate the following bioactive
substances from the extracts of the above cultures:
1. Rutin from the callus culture extract of meadowsweet
(Filipendula ulmaria) [76];
2. Quercetin from the suspension culture extract of
ginkgo (Ginkgo biloba) [77];
3. Kaempferol from the suspension culture extract of
ginkgo (G. biloba) [77];
4. Baicalin from the root culture extract of Baikal
skullcap (Scutellaria baicalensis) [78];
5. Trans-cinnamic acid from the root culture extract of
Baikal skullcap (S. baicalensis) [79];
6. Chlorogenic acid from the callus culture extract of red
clover (Trifolium pratense) [80];
7. Biochanin A from the callus culture extract of red
clover (T. pratense) [80];
8. Naringenin from the callus culture extract of alfalfa
(Medicágo satíva) [81];
9. Ursolic acid from the callus culture extract of thyme
(Thymus vulgaris) [82]; and
10. Magniferin from the root culture extract of
sweetvetch (Hedysarum neglectum) [83].
To isolate rutin from the callus culture extract of meadowsweet, the plant’s ethanol extract was evaporated under vacuum at a temperature under 40°C on an IKA RV 8 rotary evaporator (IKA, Germany). After adding deionized water to 1/4 of the concentrate’s initial volume, evaporation continued until a thick precipitate was formed. The precipitate was treated with a chloroform:ethylacetate mixture for 5 min with vigorous stirring in triplicate. The extracts were combined and mixed with anhydrous sodium sulfate (2.0 g per 100 mL of extract). The mixture was kept for 3 h at +4°C and then filtered. The residue containing a flavonoid fraction was dissolved in 50% ethanol. Then, 50.0 g of activated carbon was added to the mixture and evaporated until a dry residue was formed. The adsorbent with the extract residue was transferred to a shot filter and successively eluted with methanol, water, 7% aqueous phenol, and 15% phenol in methyl alcohol. The fraction extracted with 7% aqueous phenol was treated with 100 mL of diethyl ether in triplicate. The resulting extract was evaporated under vacuum to a thick precipitate, which was then mixed with 40.0 g of silica gel (column chromatography grade, Sigma), dried completely, and transferred to a column (5×6 cm BioRad) as a suspension in chloroform. The substances were eluted with a mixture of chloroform:ethanol (80:20) and evaporated to isolate rutin.
To isolate quercetin and kaempferol from the suspension culture extract of ginkgo, the extract was filtered through cellulose filters, diluted with water, and kept at +4°C for 48 h to filter lipid precipitates. The extract was then concentrated in a vacuum evaporator in the presence of sodium chloride (up to 10% by salt content in solution). Resinous substances were removed by decantation. Lipophilic substances were purified by liquid-liquid extraction with n-heptane to isolate terpenolactones. The aqueous phase was extracted with n-butanol in triplicate.
The three phases were combined and concentrated under vacuum until a dry precipitate was formed. The precipitate was dissolved in a water-alcohol solution and purified by liquid-liquid extraction with ethyl acetate. The resulting phase was washed with a sodium chloride solution and evaporated. The dry residue was dissolved in acetone containing 40 wt.% of water, cooled to 10°C, and filtered. Flavonogicosides were chromatographed on polyamide (Sigma) packed in a 5.3×250 mm chromatographic column on a BioLogic low-pressure chromatograph (BioRad) using gradient elution mixtures: chloroform-methanol (100:0 → 60:40) and then water-ethanol (100:0 → 0:100). The components were separated and purified by silica gel rechromatography (Lachema) using an eluent mixture of chloroform:petroleum ether (30:70), followed by recrystallization to isolate quercetin and kaempferol.
Baicalin and trans-cinnamic acid were isolated from the root culture extract of Baikal skullcap by evaporating the extract under vacuum at a temperature under 50°C. The evaporated residue was treated with diethyl ether in triplicate. The resulting ether fraction was chromatographed on silica gel (mobile phase) in a n-hexane-acetone gradient (1:0 → 0:1) to isolate flavonoids and hydroxycinnamic acids. Baicalin and trans-cinnamic acid were isolated by subsequent rechromatography on silica gel (mobile phase) with n-hexane-chloroform (1:0 → 0:1).
To isolate biochanin A from the callus culture extract of red clover, the ethanol extract was vacuumevaporated on a rotary evaporator at under 50°C. Deionized water was added to the precipitate up to 1/4 of the concentrate’s initial volume to continue evaporation to a thick precipitate. The precipitate was treated with n-hexane for 5 min in triplicate and the suspension was treated ultrasonically. The extracts were filtered through filter paper and combined. Then, they were evaporated under vacuum to a thick precipitate. The precipitate was mixed with 50.0 g of silica gel, dried, and transferred to a column (5×6 cm BioRad). Then, it was eluted with a petroleum ether-ethanol mixture (99:1, 98:2, 97:3, 95:5, 93:7, 80:20). Biochanin A was isolated from the evaporated eluates.
To isolate chlorogenic acid from the callus culture extract of red clover, the thick precipitate obtained as described above was treated with diethyl ether to isolate hydroxycinnamic and coumaric acids. The mixture was then evaporated to a dry residue and separated on silica gel (column chromatography grade, Sigma) on a column (0.65×10 cm BioRad). Then, it was eluted with isopropyl alcohol:acetic acid:hexane (65:12:23) to isolate chlorogenic acid.
Naringenin was isolated from the alfalfa extract as follows. The ethanol extract was evaporated under vacuum at under 55°C on a rotary evaporator. The residue was mixed with deionized water added to 1/4 of the concentrate’s initial volume to continue vacuum evaporation to a thick precipitate. The precipitate was placed on a 5×6 cm BioRad chromatographic column and eluted with n-hexane to collect 1-mL fractions. The resulting extracts were evaporated to a thick precipitate, which was then dissolved in ethanol and fractionated on LH-20 Sephadex (Aldrich) in toluene. Silica gel was eluted with isopropyl alcohol:water (40:60) and then evaporated under vacuum to thick residue. The residue was dissolved in ethanol and fractionated on LH-20 Sephadex (Aldrich) in a methanol gradient of 10 → 90% to isolate naringenin.
Ursolic acid was isolated from the callus culture extract of thyme. For this, the ethanolic extract was evaporated under vacuum at under 40°C on a rotary evaporator. The residue was mixed with deionized water added to 1/4 of the concentrate’s initial volume to continue vacuum evaporation to a thick precipitate. The resulting precipitate was treated three times with dichloromethane for 5 min with vigorous stirring. The extracts were combined and mixed with anhydrous sodium sulfate (20.0 g per liter of extract). The mixture was kept for 3 h and filtered through a paper filter. The filtered precipitate was dissolved in ethanol. The ethanol fraction was passed through an AN-1 anion exchanger (State Standard 20301-74) and washed with waterethanol eluents (up to 50% of ethanol). Then, it was desorbed with 0.1 M hydrochloric acid to isolate ursolic acid.
Magniferin was isolated from the root culture extract of sweetvetch. The ethanol extract was evaporated in a vacuum evaporator at 45°C. The residue was fractionated on a BioLogic low-pressure chromatograph (BioRad) using silica gel (column chromatography grade, Sigma). Petroleum ether-ethyl acetate was used as an eluent (100:0; 50:1; 20:1; 10:1; 5:1; 2:1; 1:1; and 0:1). Methanol was fed to the column to desorb gallic acid, resulting in nine 300-mL fractions collected. A crude crystal of magniferin was obtained from fraction 3. Then, it was recrystallized from a mixture of petroleum ether:acetone (20:1) and purified by rechromatography on CL6B Sepharose (Sigma – Aldrich) using a BioLogic low-pressure chromatograph (BioRad) to isolate pure mangiferin.
All the isolated bioactive substances were at least 95% pure. Their IR spectra were registered on an SF-2000 instrument (OKB Spektr, Russia).
Further, we analyzed the effect of bioactive substance concentrations on the lifespan of wild-type Caenorhabditis elegans nematodes (strain N2 Bristol, www.wormbook.org). Our study consisted of five stages described below.
Cultivation of nematodes on solid agar. Obtaining an Escherichia coli bacterial culture. E. coli OP50 was seeded on Petri dishes with a Lysogeny broth (L-broth) solid medium (HiMedia Laboratories, India). Then, under sterile conditions, one bacterial colony was selected and placed in 5–10 mL of L-broth (HiMedia Laboratories, India) to incubate at 37°C overnight with vigorous stirring. After that, the culture was transferred to a refrigerator and stored at +4°C.
Inoculating E. coli OP50 on NGM agar plates. 50 μL of the E. coli OP50 overnight culture was placed in the center of a 100-mm Petri dish. Using a sterile glass rod, the drop was distributed over the center of the dish in the shape of a square, without touching the walls, and incubated at 37°C for a day. After incubation, the dishes were wrapped in parafilm and stored in the refrigerator for several weeks.
Preparing NGM agar plates. After autoclaving, the sterile NGM agar was cooled to 55°C in a water bath. Then, the cooled nutrient medium was mixed with 1 mL of 1 M CaCl2, 1 mL of 5 mg/mL cholesterol in alcohol, 1 mL of 1 M MgSO4, and 25 mL of 1 M K3PO4 buffer. After thorough mixing, it was poured into sterile Petri dishes, 20 mL each. To ensure the absence of bacterial contamination, the dishes were left for 2–3 days at room temperature.
Transferring nematodes to new NGM agar dishes. The nematodes were transferred in two ways: with a loop and with a piece of agar. The first method involved hooking a nematode with a calcined and cooled bacteriological loop and planting it on a bacterial lawn in the center of a new NGM Petri dish with agar. The second method involved cutting a 0.5×0.5 cm piece of agar containing a nematode with a sterile scalpel and transferring it to the center of the dish surface down. The dishes were incubated at 20°C.
Nematode synchronization. 5–10 mL of sterile water was pipetted on the surface of the dish containing a nematode until its eggs were completely attached to the agar. The liquid from the Petri dish was placed in a 50 mL tube and centrifuged for 2 min (1200 rpm). Then, the supernatant was removed and the precipitate was washed with 10 mL of distilled water to repeat centrifugation under the above conditions.
After repeated centrifugation, the supernatant was removed and the precipitate was mixed with 5 mL of a freshly prepared mixture of 1 mL of 10 N NaOH, 2.5 mL of household bleach, and 6.5 mL of H2O. The mixture was thoroughly vortexed (Biosan, Latvia) for 10 min with 2 min intervals to observe the hydrolysis of nematodes under an Axio Observer Z1 microscope (Karl Zeiss, Germany). At the end of the process, 5 mL of M9 medium was added to neutralize the reaction. The resulting mixture was centrifuged for 2 min (2500 rpm). After that, the supernatant was removed and the precipitate was mixed with 10 mL of sterile water to repeat the washing and centrifugation three times. Then, the precipitate was washed with 10 mL of S-medium and the supernatant was removed. Finally, 10 mL of S-medium was added and the test tube with nematode eggs was placed on a slow shaker for a day at room temperature for the complete transfer of the nematodes to the L1 stage.
Cultivation of nematodes in a liquid medium. After the nematodes passed to the L1 stage, an overnight bacterial culture of E. coli OP50 was added to the S-medium. The culture had previously been washed and resuspended in the S-medium to a bacterial concentration of 0.5 mg/mL. Then, 120 μL amounts of the suspension containing the bacteria and nematodes were poured into each well of a 96-well plate (TPP, Switzerland). The plate was sealed with a film and left for 48 h at 20°C.
After that, 15 μL of 1.2 mM 5-fluoro-2-deoxyuredin was poured into each well of the plate to prevent the nematodes from reproduction and left for a day at 20°C. At the end of incubation, the nematodes entered the L4 stage. Then, 15-μL amounts of the bioactive substances under study were added to the wells in accordance with the experiment plan.
Preparation of bioactive substances. Stock solutions of bioactive substances were prepared in dimethyl sulfoxide at a concentration of 10 mmol/L. The substances were tested by diluting stock solutions in sterile distilled water to concentrations of 2000, 1000, 500, and 100 μM. Each well was filled with 15 μL of freshly prepared stock solutions so that working concentrations of each bioactive substance reached 2000, 1000, 500, and 100 μmol/L, respectively. The stocks were stored at 4°C.
Effects of bioactive substances on nematode lifespan. The effect of bioactive substances at concentrations of 0, 10, 50, 100, and 200 μmol/L on the lifespan of C. elegans was determined by the number of nematodes surviving in the presence of the tested substances. The experiment was carried out in 6-fold repetitions using 96-well plates and a liquid S-medium for nematode cultivation. The numbers of live and dead nematodes were counted every 4–7 days during the 61-day experiment. The experiment was considered completed when there were no live nematodes left in the control group.
Each concentration of bioactive substances was studied in 6-fold repetitions. Statistical data were analyzed in the Microsoft Office Excel 2007. Statistical analysis was performed using a paired Student’s t-test for each pair of interests. Differences were considered statistically significant at p < 0.05.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
The effects of the bioactive substances obtained from the extracts of suspension, callus, and root cultures of Siberian medicinal plants on the lifespan of Caenorhabditis elegans L. nematodes are graphically presented in Fig. 1.
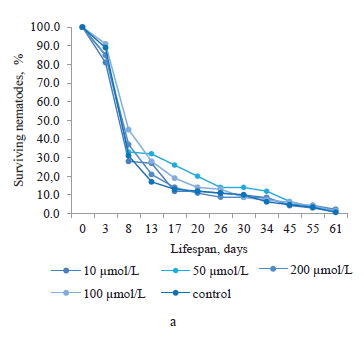
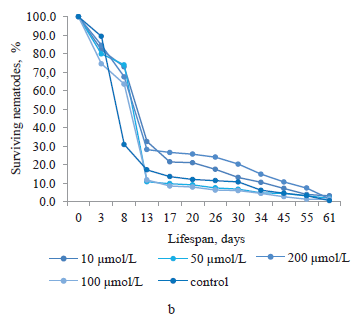




Quercetin, which was obtained from the suspension culture extract of ginkgo, had a significant effect on the lifespan of C. elegans nematodes at concentrations of 10 and 100 μmol/L (Fig. 1b). The proportion of surviving nematodes was 32.6–4.6% from day 8 to day 45 of the experiment. On day 8, all the concentrations of quercetin had a positive effect on the lifespan. The proportions of surviving nematodes treated with 10, 50, 100 and 200 μmol/L of this bioactive substance were 72.9, 74.0, 67.5, and 63.6%, respectively (higher than in the control nematodes by 41.9, 43.0, 36.5, and 32.6%, respectively).
Of special interest was kaempferol obtained from the suspension culture extract of ginkgo (Fig. 1c). At a concentration of 50 μmol/l, this substance increased the lifespan of nematodes throughout the experiment (except for 3 days), compared to the control. The nematode population was active, reaching 10.3% on day 61. We also observed kaempferol’s positive effect at a concentration of 10 μmol/L in the period of 8 to 61 days. The maximum proportion of surviving nematodes treated with this concentration was registered on day 8 at 48.6%, which was 17.6% higher than in the control group. However, the greatest increase in the nematode lifespan was provided by a concentration of 200 μmol/L on day 8, with the survival rate of 61.6% (by 30.6% higher than in the control nematodes).
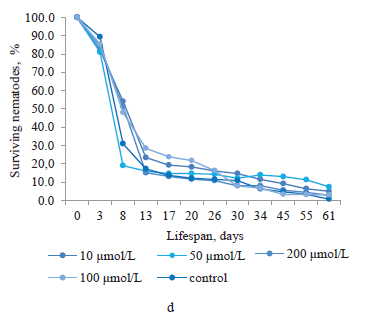
Baicalin was produced from the root culture of Baikal skullcap (Fig. 1d). At concentrations of 10, 100, and 200 μmol/L, it increased the lifespan of nematodes from day 8 to day 13. After that period, the number of surviving nematodes exposed to 200 μmol/L baicalin became lower than in the control group. During days 13–17, their lifespan increased only at baicalin’s concentrations of 10 and 100 μmol/L. From day 17 until the end of the experiment, the number of surviving nematodes treated with 10, 50, and 200 μmol/L baicalin was greater than in the control group. Noteworthily, the end of the experiment saw greater proportions of surviving nematodes treated with baicalin at all concentrations (10, 50, 100, and 200 μmol/L) than that of the control (by 4.3, 6.7, 2.3, and 2.1%, respectively).
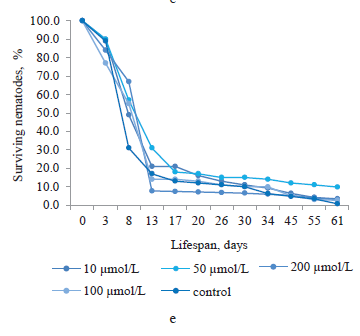
Trans-cinnamic acid was isolated from the root culture extract of Baikal skullcap (Fig. 1e). As we can see, on day 8, its concentrations of 10, 50, 100, and 200 μmol/L increased the lifespan of nematodes by 18.1, 26.3, 24.1, and 36.6%, respectively. During days 13–34, the concentration of 200 μmol/L had no positive effect on the lifespan of nematodes, unlike the other concentrations. However, from day 34 to the end of the experiment, trans-cinnamic acid at all concentrations increased the percentage of surviving nematodes. The greatest increase in the lifespan was observed in the nematodes treated with 50 μmol/L of this bioactive substance (9.8%).
Chlorogenic acid obtained from the callus culture extract of red clover showed a generally positive effect on the lifespan of C. elegans (Fig. 1f). As can be seen, 100 μmol/L of this substance increased the survival of nematodes throughout the experiment, with the greatest increase (by 40.1%) on day 8. The other concentrations showed varying survival rates. Days 8–13 saw greater lifespans in the nematodes exposed to chlorogenic acid at all four concentrations. During days 13–26, increased lifespan was provided by the concentrations of 10, 50, and 100 μmol/L. The maximum survival rate was observed on day 20 (27.7%) in the nematodes treated with 100 μmol/L of chlorogenic acid. From day 26 to day 34, the concentration of 200 μmol/L had no effect on the lifespan of C. elegans. However, from day 45 to the end of the experiment, chlorogenic acid had a positive effect again at all its concentrations.
Figure 1g shows the effect of biochanin A isolated from red clover callus culture. As we can see, the best survival rate was provided by this substance at 100 μmol/L. Unlike the other concentrations, this concentration had a positive effect on C. elegans throughout the entire experiment. Days 8–20 saw the highest survival rates, with the greatest increase in lifespan occurring on day 13 (by 45.2% compared to the control). Noteworthily, 200 μmol/L of biochanin A had a negative effect on the survival and lifespan of C. elegans almost throughout the experiment, except for the very end. On day 61, the proportion of surviving nematodes increased by 1.8% and amounted to 2.5% (compared to 0.7% in the control group). The concentrations of 10 and 50 μmol/L increased the survival of nematodes during days 13–45 by an average of 8.4 and 9.1%, respectively. Both concentrations provided maximum lifespan increases during days 13–20 and had a weaker effect towards the end of the experiment. From day 26 to day 61, biochanin A at 10, 50, and 200 μmol/L had no significant effect on the lifespan of nematodes compared to the control group.
Naringenin was isolated from the extract of alfalfa callus culture. As shown by Fig. 1h, its concentration of 100 μmol/L had the greatest effect on the lifespan of nematodes compared to the other concentrations, especially during days 8–26. During that period, the survival of nematodes increased by an average of 27.3% compared to the control group, with maximum survival on day 13 (by 35.4%). The other concentrations of naringenin (10, 50, and 200 μmol/L) did not have a significant effect on the survival or lifespan of nematodes.
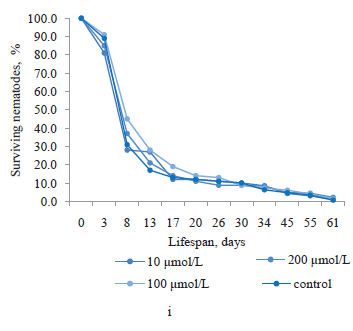
Ursolic acid was isolated from thyme callus culture. According to Fig. 1i, it had no significant effect on the lifespan of nematodes at all its concentrations. The greatest increase in survival (by 14.1%) was observed on day 8 in the nematodes treated with 100 μmol/L of ursolic acid.
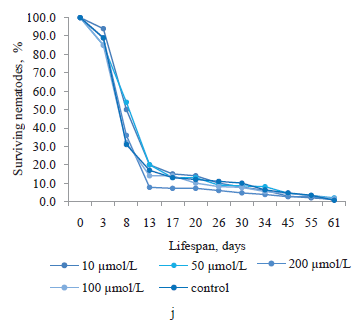
Similarly, we found no positive effect in magniferin obtained from the root culture extract of sweetvetch (Fig. 1j). Moreover, its concentrations of 100 and 200 μmol/L reduced the proportion of surviving nematodes from day 13 to day 55 of the experiment. The greatest increases in the lifespan of nematodes were observed at magniferin concentrations of 10 and 50 μmol/L on day 8, amounting to 19.3 and 24.2%, respectively. At the end of the experiment, the longest lifespan was demonstrated by the nematodes exposed to 100 μmol/L of magniferin (1.4%).
ВЫВОДЫ
Having applied HPLC methods, we isolated the following bioactive substances from the extracts of callus, suspension, and root cultures of medicinal plants growing in the Siberian Federal Okrug: rutin – from the callus culture extract of meadowsweet (Filipendula ulmaria L.); quercetin – from the suspension culture extract of ginkgo (Ginkgo biloba L.); kaempferol – from the suspension culture extract of ginkgo (G. biloba); baicalin – from the root culture extract of Baikal skullcap (Scutellaria baicalensis L.); trans-cinnamic acid – from the root culture extract of Baikal skullcap (S. baicalensis); chlorogenic acid – from the callus culture extract of red clover (Trifolium pretense L.); biochanin A – from the callus culture extract of red clover (T. pratense); naringenin – from the callus culture extract of alfalfa (Medicágo sativa L.); ursolic acid – from the callus culture extract of thyme (Thymus vulgaris L.); and magniferin – from the root culture extract of sweetvetch (Hedysarum neglectum L.). All the bioactive substances were at least 95% pure and were registered using IR spectroscopy on an SF-2000 instrument (OKB Spektr, Russia).
We determined the effect of the above bioactive substances at concentrations of 10, 50, 100, and 200 μmol/L on the lifespan of Caenorhabditis elegans nematodes, which are widely used as model organisms to study the aging process. We used 96-well plates for the experiment that lasted 61 days. Surviving nematodes were counted every 4–7 days and the experiment was considered completed when there were no live nematodes left in the control group. Stock solutions of the following bioactive substances were prepared for the experiment: rutin, quercetin, kaempferol, baicalin, trans-cinnamic acid, chlorogenic acid, biochanin A, naringenin, ursolic acid, and magniferin.
Trans-cinnamic acid, baicalin, rutin, ursolic acid, and magniferin did not significantly increase the lifespan of the nematodes.
Chlorogenic acid and naringenin had a little effect on the lifespan of nematodes, while quercetin, kaempferol, and biochanin A demonstrated their high survival.
Noteworthily, the greatest proportions of surviving nematodes treated with various concentrations of bioactive substances were recorded on days 8 to 13 for all the experimental samples. Then, the lifespan of C. elegans decreased and their survival rates approached those of the control group. Thus, 200 μmol/L of kaempferol, 10 μmol/L of quercetin (both obtained from ginkgo suspension culture extract), and 100 μmol/L of biochanin A (obtained from red clover callus culture extract) increased the lifespan of C. elegans nematodes by 30.6, 41.9, and 45.2%, respectively, compared to the control (days 8 and 13). These results suggest that the mentioned bioactive substances can be effectively used as anti-aging agents.
Вклад авторов
All the authors are equally responsible for the research results and the manuscript.КОНФЛИКТ ИНТЕРЕСОВ
The authors declare no conflict of interest.ФИНАНСИРОВАНИЕ
The study was financed by the Ministry of Science and Higher Education of the Russian Federation (Minobrnauka) (project FZSR-2020-0006 “Screening bioactive plant-based substances with geroprotective properties and developing technology for producing anti-aging nutraceuticals”).СПИСОК ЛИТЕРАТУРЫ
- WHO: People living longer and healthier lives but COVID-19 threatens to throw progress off track [Internet]. [cited 2022 Jan 15]. Available from: https://www.who.int/news/item/13-05-2020-people-living-longer-and-healthier-lives-but-covid-19-threatens-to-throw-progress-off-track
- Bosch-Morell F, Villagrasa V, Ortega T, Acero N, Muñoz-Mingarro D, González-Rosende M, et al. Medicinal plants and natural products as neuroprotective agents in age–related macular degeneration. Neural Regeneration Research. 2020;15(12):2207–2216. https://doi.org/10.4103/1673-5374.284978
- Elfawy HA, Das B. Crosstalk between mitochondrial dysfunction, oxidative stress, and age related neurodegenerative disease: Etiologies and therapeutic strategies. Life Sciences. 2019;218:165–184. https://doi.org/10.1016/j.lfs.2018.12.029
- Tan BL, Norhaizan ME, Liew W-P-P, Rahman HS. Antioxidant and oxidative stress: A mutual interplay in age–related diseases. Frontiers in Pharmacology. 2018;9. https://doi.org/10.3389/fphar.2018.01162
- Prosekov AYu, Dyshlyuk LS, Milentyeva IS, Sykhikh SA, Babich OO, Ivanova SA, et al. Antioxidant and antimicrobial activity of bacteriocin-producing strains of lactic acid bacteria isolated from the human gastrointestinal tract. Progress in Nutrition. 2017;19(1):67–80. https://doi.org/10.23751/pn.v19i1.5147
- Harman D. Aging: A theory based on free radical and radiation chemistry. Journal of Gerontology. 1956;11(3):298–300. https://doi.org/10.1093/geronj/11.3.298
- McGuire PJ. Mitochondrial dysfunction and the aging immune system. Biology. 2019;8(2). https://doi.org/10.3390/biology8020026
- Dębowska K, Smulik-Izydorczyk R, Pięta J, Adamus J, Michalski R, Sikora A. Oxidation of the selected probes for detection of reactive oxygen species (ROS) and reactive nitrogen species (RNS) in aqueous solutions of nitric oxide donors. Free Radical Biology and Medicine. 2018;120. https://doi.org/10.1016/j.freeradbiomed.2018.04.258
- Jakubczyk K, Dec K, Kałduńska J, Kawczuga D, Kochman J, Janda, K. Reactive oxygen species – sources, functions, oxidative damage. Polski merkuriusz lekarski: organ Polskiego Towarzystwa Lekarskiego. 2020;48(284):124–127.
- He L, He T, Farrar S, Ji L, Liu T, Ma X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cellular Physiology and Biochemistry. 2017;44(2):532–553. https://doi.org/10.1159/000485089
- van de Lagemaat EE, de Groot LCPGM, van den Heuvel EGHM. Vitamin B12 in relation to oxidative stress: A systematic review. Nutrients. 2019;11(2). https://doi.org/10.3390/nu11020482
- Singh A, Kukreti R, Saso L, Kukreti S. Oxidative stress: A key modulator in neurodegenerative diseases. Molecules. 2019;24(8). https://doi.org/10.3390/molecules24081583
- Warraich UA, Hussain F, Kayani HUR. Aging – Oxidative stress, antioxidants and computational modeling. Heliyon. 2020;6(5). https://doi.org/10.1016/j.heliyon.2020.e04107
- Mirończuk-Chodakowska I, Witkowska AM, Zujko ME. Endogenous non-enzymatic antioxidants in the human body. Advances in Medical Sciences. 2018;63(1):68–78. https://doi.org/10.1016/j.advms.2017.05.005
- Santos AL, Sinha S, Lindner AB. The good, the bad, and the ugly of ROS: New insights on aging and aging–related diseases from eukaryotic and prokaryotic model organisms. Oxidative Medicine and Cellular Longevity. 2018;2018. https://doi.org/10.1155/2018/1941285
- Liu R, Mabury SA. Synthetic phenolic antioxidants: A review of environmental occurrence, fate, human exposure, and toxicity. Environmental Science and Technology. 2020;54(19):11706–11719. https://doi.org/10.1021/acs.est.0c05077
- Asyakina LK, Fotina NV, Izgarysheva NV, Slavyanskiy AA, Neverova OA. Geroprotective potential of in vitro bioactive compounds isolated from yarrow (Achilleae millefolii L.) cell cultures. Foods and Raw Materials. 2021;9(1):126–134. https://doi.org/10.21603/2308-4057-2021-1-126-134
- Gadouche L, Zidane A, Zerrouki K, Azouni K, Bouinoune S. Cytotoxic effect of Myrtus communis, Aristolochia longa, and Calycotome spinosa on human erythrocyte cells. Foods and Raw Materials. 2021;9(2):379–386. https://doi.org/10.21603/2308-4057-2021-2-379-386
- Hassan W, Noreen H, Rehman S, Gul S, Amjad Kamal M, Kamdem JP, et al. Oxidative stress and antioxidant potential of one hundred medicinal plants. Current Topics in Medicinal Chemistry. 2017;17(12):1336–1370. https://doi.org/10.2174/1568026617666170102125648
- Song R, Wu Q, Yun Z, Zhao L. Advances in antioxidative bioactive macromolecules. IOP Conference Series: Earth and Environmental Science. 2020;512(1). https://doi.org/10.1088/1755-1315/512/1/012094
- Nascimento-Souza MA, de Paiva PG, Pérez-Jiménez J, do Carmo Castro Franceschini S, Ribeiro AQ. Estimated dietary intake and major food sources of polyphenols in elderly of Viçosa, Brazil: a population-based study. European Journal of Nutrition. 2018;57(2):617–627. https://doi.org/10.1007/s00394-016-1348-0
- Fraga CG, Croft KD, Kennedy DO, Tomás-Barberán FA. The effects of polyphenols and other bioactives on human health. Food and Function. 2019;10(2):514–528. https://doi.org/10.1039/C8FO01997E
- Majidinia M, Bishayee A, Yousefi B. Polyphenols: Major regulators of key components of DNA damage response in cancer. DNA Repair. 2019;82. https://doi.org/10.1016/j.dnarep.2019.102679
- Abdel‐Moneim AME, Shehata AM, Alzahrani SO, Shafi ME, Mesalam NM, Taha AE, et al. The role of polyphenols in poultry nutrition. Journal of Animal Physiology and Animal Nutrition. 2020;104(6):1851–1866. https://doi.org/10.1111/jpn.13455
- Tungmunnithum D, Thongboonyou A, Pholboon A, Yangsabai A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines. 2018;5(3). https://doi.org/10.3390/medicines5030093
- Papaevgeniou N, Chondrogianni N. Anti-aging and anti-aggregation properties of polyphenolic compounds in C. elegans. Current Pharmaceutical Design. 2018;24(19):2107–2120. https://doi.org/10.2174/1381612824666180515145652
- Chaudhary MK, Rizvi SI. Invertebrate and vertebrate models in aging research. Biomedical Papers. 2019;163(2):114–121. https://doi.org/10.5507/bp.2019.003
- Ye Y, Gu Q, Sun X. Potential of Caenorhabditis elegans as an antiaging evaluation model for dietary phytochemicals: A review. Comprehensive Reviews in Food Science and Food Safety. 2020;19(6):3084–3105. https://doi.org/10.1111/1541-4337.12654
- Reiss AP, Rankin CH. Gaining an understanding of behavioral genetics through studies of foraging in Drosophila and learning in C. elegans. Journal of Neurogenetics. 2021;35(3):119–131. https://doi.org/10.1080/01677063.2021.1928113
- Douglas AE. The Drosophila model for microbiome research. Lab Animal. 2018;47(6):157–164. https://doi.org/10.1038/s41684-018-0065-0
- Markaki M, Tavernarakis N. Caenorhabditis elegans as a model system for human diseases. Current Opinion in Biotechnology. 2020;63:118–125. https://doi.org/10.1016/j.copbio.2019.12.011
- Mack HID, Heimbucher T, Murphy CT. The nematode Caenorhabditis elegans as a model for aging research. Drug Discovery Today: Disease Models. 2018;27:3–13. https://doi.org/10.1016/j.ddmod.2018.11.001
- Salzer L, Witting M. Quo vadis Caenorhabditis elegans metabolomics – A review of current methods and applications to explore metabolism in the nematode. Metabolites. 2021;11(5). https://doi.org/10.3390/metabo11050284
- Martel J, Wu C-Y, Peng H-H, Ko Y-F, Yang, H-C, Young, JD, et al. Plant and fungal products that extend lifespan in Caenorhabditis elegans. Microbial Cell. 2020;7(10):255–269. https://doi.org/10.15698/mic2020.10.731
- Elkabti AB, Issi L, Rao RP. Caenorhabditis elegans as a model host to monitor the Candida infection processes. Journal of Fungi. 2018;4(4). https://doi.org/10.3390/jof4040123
- Ayuda-Durán B, González-Manzano S, González-Paramás AM, Santos-Buelga C. Caernohabditis elegans as a model organism to evaluate the antioxidant effects of phytochemicals. Molecules. 2020;25(14). https://doi.org/10.3390/molecules25143194
- Van Pelt KM, Truttmann MC. Caenorhabditis elegans as a model system for studying aging-associated neurodegenerative diseases. Translational Medicine of Aging. 2020;4:60–72. https://doi.org/10.1016/j.tma.2020.05.001
- Bouyanfif A, Jayarathne S, Koboziev I, Moustaid-Moussa N. The nematode Caenorhabditis elegans as a model organism to study metabolic effects of ω-3 polyunsaturated fatty acids in obesity. Advances in Nutrition. 2019;10(1):165–178. https://doi.org/10.1093/advances/nmy059
- Wang C, Xia C, Zhu Y, Zhang H. Innovative fluorescent probes for in vivo visualization of biomolecules in living Caenorhabditis elegans. Cytometry Part A. 2021;99(6):560–574. https://doi.org/10.1002/cyto.a.24325
- Sandhof CA, Hoppe SO, Tittelmeier J, Nussbaum-Krammer C. C. elegans models to study the propagation of prions and prion-like proteins. Biomolecules. 2020;10(8). https://doi.org/10.3390/biom10081188
- Hunt PR. The C. elegans model in toxicity testing. Journal of Applied Toxicology. 2017;37(1):50–59. https://doi.org/10.1002/jat.3357
- Martel J, Wu C-Y, Peng H-H, Ko Y-F, Yang H-C, Young JD, et al. Plant and fungal products that extend lifespan in Caenorhabditis elegans. Microbial Cell. 2020;7(10):255–269. https://doi.org/10.15698/mic2020.10.731
- Backes C, Martinez-Martinez D, Cabreiro F. C. elegans: A biosensor for host-microbe interactions. Lab Animal. 2021;50(5):127–135. https://doi.org/10.1038/s41684-021-00724-z
- Ma L, Zhao Y, Chen Y, Cheng B, Peng A, Huang K. Caenorhabditis elegans as a model system for target identification and drug screening against neurodegenerative diseases. European Journal of Pharmacology. 2018;819:169–180. https://doi.org/10.1016/j.ejphar.2017.11.051
- Liang JJH, McKinnon IA, Rankin CH. The contribution of C. elegans neurogenetics to understanding neurodegenerative diseases. Journal of Neurogenetics. 2020;34(3–4):527–548. https://doi.org/10.1080/01677063.2020.1803302
- Keith Blackwell T, Sewell AK, Wu Z, Han M. TOR signaling in Caenorhabditis elegans development, metabolism, and aging. Genetics. 2019;213(2):329–360. https://doi.org/10.1534/genetics.119.302504
- Fedorova AM, Dmitrieva AI, Dyshlyuk LS. Cultivation of wild medicinal plants of the SFD in vitro to accumulate the potential geroprotectors. Scientific Works of the North Caucasus Federal Scientific Center of Horticulture, Viticulture, Winemaking. 2020;30:134–138. (In Russ.). https://doi.org/10.30679/2587-9847-2020-30-134-138
- Baranenko D, Bespalov V, Nadtochii L, Shestopalova I, Chechetkina A, Lepeshkin A, et al. Development of encapsulated extracts on the basis of meadowsweet (Filipendula ulmaria) in the composition of functional foods with oncoprotective properties. 2019;17(5):1829–1838. https://doi.org/10.15159/ar.19.155
- Bespalov VG, Alexandrov VA, Vysochina GI, Kostikova VA, Semenov AL, Baranenko DA. Inhibitory effect of Filipendula ulmaria on mammary carcinogenesis induced by local administration of methylnitrosourea to target organ in rats. Anti-Cancer Agents in Medicinal Chemistry. 2018;18(8):1177–1183. https://doi.org/10.2174/1871520618666180402125913
- Shaldayeva TM, Vysochina GI, Kostikova VA. Phenolic compounds and antioxidant activity of some species of the genus Filipendula Mill. (Rosaceae). Proceedings of Voronezh State University. Series: Chemistry. Biology. Pharmacy. 2018;(1):204–212. (In Russ.).
- Kurkin VA, Sazanova KN, Zaitseva EN, Sharipova SKh, Pravdivtseva OE, Avdeeva EV, et al. Antidepressant activity of flavonoids and thick extract from Filipendula ulmaria fruit. Pharmaceutical Chemistry Journal. 2020;54(8):797–799. https://doi.org/10.1007/s11094-020-02276-x
- Adamczak A, Ożarowski M, Karpiński TM. Antibacterial activity of some flavonoids and organic acids widely distributed in plants. Journal of Clinical Medicine. 2020;9(1). https://doi.org/10.3390/jcm9010109
- Satari A, Ghasemi S, Habtemariam S, Asgharian S, Lorigooini Z. Rutin: A flavonoid as an effective sensitizer for anticancer therapy; insights into multifaceted mechanisms and applicability for combination therapy. Evidence-based Complementary and Alternative Medicine. 2021;2021. https://doi.org/10.1155/2021/9913179
- Wang C, Shang S, Zheng X, Lei P, Han J, Yuan L, et al. Fluorescent sensors based on Cu-doped carbon quantum dots for the detection of rutin. Journal of the Brazilian Chemical Society. 2019;30(5):988–996. https://doi.org/10.21577/0103-5053.20180245
- Negahdari R, Bohlouli S, Sharifi S, Maleki Dizaj S, Rahbar Saadat Y, Khezri K, et al. Therapeutic benefits of rutin and its nanoformulations. Phytotherapy Research. 2021;35(4):1719–1738. https://doi.org/10.1002/ptr.6904
- Singh SK, Srivastav S, Castellani RJ, Plascencia-Villa G, Perry G. Neuroprotective and antioxidant effect of Ginkgo biloba extract against AD and other neurological disorders. Neurotherapeutics. 2019;16(3):666–674. https://doi.org/10.1007/s13311-019-00767-8
- Priyanka A, Sindhu G, Shyni GL, Preetha Rani MR, Nisha VM, Raghu KG. Bilobalide abates inflammation, insulin resistance and secretion of angiogenic factors induced by hypoxia in 3T3-L1 adipocytes by controlling NF-κB and JNK activation. International Immunopharmacology. 2017;42:209–217. https://doi.org/10.1016/j.intimp.2016.11.019
- Eisvand F, Razavi BM, Hosseinzadeh H. The effects of Ginkgo biloba on metabolic syndrome: A review. Phytotherapy Research. 2020;34(8):1798–1811. https://doi.org/10.1002/ptr.6646
- Patel RV, Mistry BM, Shinde SK, Syed R, Singh V, Shin H-S. Therapeutic potential of quercetin as a cardiovascular agent. European Journal of Medicinal Chemistry. 2018;155:889–904. https://doi.org/10.1016/j.ejmech.2018.06.053
- Li X, Zhou N, Wang J, Liu Z, Wang X, Zhang Q, et al. Quercetin suppresses breast cancer stem cells (CD44+/CD24−) by inhibiting the PI3K/Akt/mTOR-signaling pathway. Life Sciences. 2018;196:56–62. https://doi.org/10.1016/j.lfs.2018.01.014
- Bahadır HM, Sarıgöz T, Topuz Ö, Sevim Y, Ertan T, Sarıcı İŞ. Protective effects of quercetin on hepatic ischemia-reperfusion injury. Istanbul Medical Journal. 2018;19(1):47–51. https://doi.org/10.5152/imj.2017.72325
- Kashyap D, Sharma A, Tuli HS, Sak K, Punia S, Mukherjee TK. Kaempferol – A dietary anticancer molecule with multiple mechanisms of action: Recent trends and advancements. Journal of Functional Foods. 2017;30:203–219. https://doi.org/10.1016/j.jff.2017.01.022
- Zhao T, Tang H, Xie L, Zheng Y, Ma Z, Sun Q, et al. Scutellaria baicalensis Georgi.(Lamiaceae): a review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Journal of Pharmacy and Pharmacology. 2019;71(9):1353–1369. https://doi.org/10.1111/jphp.13129
- Wang Z-L, Wang S, Kuang Y, Hu Z-M, Qiao X, Ye M. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharmaceutical Biology. 2018;56(1):465–484. https://doi.org/10.1080/13880209.2018.1492620
- Liao H, Ye J, Gao L, Liu Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomedicine and Pharmacotherapy. 2021;133. https://doi.org/10.1016/j.biopha.2020.110917
- Antonescu A-I, Miere F, Fritea L, Ganea M, Zdrinca M, Dobjanschi L, et al. Perspectives on the combined effects of Ocimum basilicum and Trifolium pratense extracts in terms of phytochemical profile and pharmacological effects. Plants. 2021;10(7). https://doi.org/10.3390/plants10071390
- Egan LM, Hofmann RW, Seguin P, Ghamkhar K, Hoyos-Villegas V. Pedigree analysis of pre-breeding efforts in Trifolium spp. germplasm in New Zealand. BMC Genet. 2020;21(1). https://doi.org/10.1186/s12863-020-00912-9
- Kubes J, Skalicky M, Tumova L, Martin J, Hejnak V, Martinkova J. Vanadium elicitation of Trifolium pretense L. cell culture and possible pathways of produced isoflavones transport across the plasma membrane. Plant Cell Reports. 2019;38(5):657–671. https://doi.org/10.1007/s00299-019-02397-y
- Wang G, Wang J, Liu W, Nisar MF, El-Esawi MA, Wan C. Biological activities and chemistry of triterpene saponins from Medicago species: An update review. Evidence-based Complementary and Alternative Medicine. 2021;2021. https://doi.org/10.1155/2021/6617916
- Sharma P, Kumar V, Guleria P. Naringin: Biosynthesis and pharmaceutical applications. Indian Journal of Pharmaceutical Sciences. 2019;81(6): 988–999. https://doi.org/10.36468/pharmaceutical-sciences.596
- Bhia M, Motallebi M, Abadi B, Zarepour A, Pereira-Silva M, Saremnejad F, et al. Naringenin nano-delivery systems and their therapeutic applications. Pharmaceutics. 2021;13(2). https://doi.org/10.3390/pharmaceutics13020291
- Salehi B, Mishra AP, Shukla I, Sharifi‐Rad M, Contreras MDM, Segura‐Carretero A, et al. Thymol, thyme, and other plant sources: Health and potential uses. Phytotherapy Research. 2018;32(9):1688–1706. https://doi.org/10.1002/ptr.6109
- Abbaszadeh S, Teimouri H, Farzan B. An ethnobotanical study of medicinal plants with antianxiety and antidepressant effects in Shahrekord. Egyptian Journal of Veterinary Sciences. 2019;50(1):81–87.
- Seo DY, Lee SR, Heo J-W, No M-H, Rhee BD, Ko KS, et al. Ursolic acid in health and disease. Korean Journal of Physiology and Pharmacology. 2018;22(3):235–248. https://doi.org/10.4196/kjpp.2018.22.3.235
- Mlala S, Oyedeji AO, Gondwe M, Oyedeji OO. Ursolic acid and its derivatives as bioactive agents. Molecules. 2019;24(15). https://doi.org/10.3390/molecules24152751
- Bijttebier S, Van Der Auwera A, Voorspoels S, Noten B, Hermans N, Pieters L, et al. A first step in the quest for the active constituents in Filipendula ulmaria (meadowsweet): Comprehensive phytochemical identification by liquid chromatography coupled to quadrupole-orbitrap mass spectrometry. Planta Medica. 2016;82(6):559–572. https://doi.org/10.1055/s-0042-101943
- Budeč M, Bošnir J, Racz A, Lasić D, Brkić D, Mosović Ćuić A, et al. Verification of authenticity of Ginkgo biloba L. Leaf extract and its products present on the Croatian market by analysis of quantity and ratio of ginkgo flavone glycosides (quercetin, kaempferol and isorhamnetin) to terpene trilactones to the effect of unmasking counterfeit drugs endangering patient health. Acta Clinica Croatica. 2019;58(4):672–692. https://doi.org/10.20471/acc.2019.58.04.15
- Boyko NN, Pisarev DI, Zhilyakova ET, Maljutina AYu, Novikov OO, Bocharnikova MA. Study of baicalin hydrolysis kinetics in the process of its extraction from Scutellaria baicalensis Georgi roots. Pharmacy and Pharmacology. 2019;7(3):129–137. (In Russ.). https://doi.org/10.19163/2307-9266-2019-7-3-129-137
- Lay H-L, Chen C-C, Chiang S-T. Simultaneous analysis of nine components in “Byi-Liang-Tang” preparation by high performance liquid chromatography. Journal of Food and Drug Analysis. 2004;12(2):115–119. https://doi.org/10.38212/2224-6614.2654
- Dyshlyuk LS, Fedorova AM, Loseva AI, Eremeeva NI. Callus cultures of Thymus vulgaris and Trifolium pratense as a source of geroprotectors. Food Processing: Techniques and Technology. 2021;51(2):423–432. https://doi.org/10.21603/2074-9414-2021-2-423-432
- Chen XM, Wang FF, Wang YQ, Li XL, Wang AR, Wang CL, et al. Discrimination of the rare medicinal plant Dendrobium officinale based on naringenin, bibenzyl, and polysaccharides. Science China Life Sciences. 2012;55(12):1092–1099. https://doi.org/10.1007/s11427-012-4419-3
- Shimada A, Ueno H, Inagaki M. Glutaminase inhibitory activities of pentacyclic triterpenes isolated from Thymus vulgaris L. Natural Product Research. 2021;36(11):1864–2868. https://doi.org/10.1080/14786419.2021.1921766
- Fedorova YuS, Sukhikh AS, Suslov NI, Zakhar YuV, Soboleva OM. Chromatography of hedysarum extracts on modified Sepharose CL sorbents. Fundamental and Clinical Medicine. 2019;4(3):61–67. (In Russ.). https://doi.org/10.23946/2500-0764-2019-4-3-61-67
