

Аннотация
Introduction. The functional basis of protopectin complex can be represented as a network of regions that consist of homogalacturonan sequences and a base of rhamnogalacturonans-I, i.e. rhamnosyl-containing branching sites. Enzymatic isolation of these regions is possible only at a certain minimal native degree of polymerization. The research objective was to develop a system of criteria for assessing the potential applicability of the enzymatic transformation of plant protopectin complex.Study objects and methods. The research featured the polymerization degree of the homogalacturonan regions of the protopectin complex and produced a system of assessment criteria for the enzymatic fragmentation potential of the protopectin complex. The theoretical calculations were based on the values of the mass fractions of rhamnosyl and galacturonide residues in plant cell walls. The result was a new polymerization degree analytical function.
Results and discussion. The ratio of the mass fractions of rhamnosyl and galacturonide residues in the water-insoluble plant tissue served as a dimensionless criterion of applicability. The rational condition for the dimensionless criterion of applicability was based on the fundamental constraint for homogalacturonan regions in the protopectin complex. It was expressed by a fundamental inequation. The rational area for determining the numerical values of the applicability criterion was presented as
Conclusion. The new approach offers an assessment of the enzymatic fragmentation potential of the plant protopectin complex by homoenzyme preparations. The approach is in fact the second stage of the decision tree in the science-based technology for pectin and its products.
Ключевые слова
Protopectin complex, rhamnogalacturonan-I, homogalacturonan, transformation, criterion assessmentВВЕДЕНИЕ
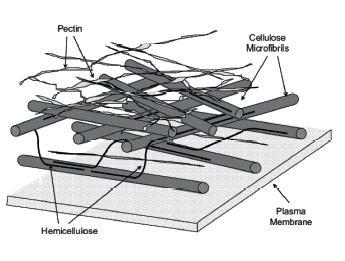
The biopolymer complex of plant tissue cell walls is a complex conglomerate of intertwined branched supramolecular networks of the protopectin complex and the hemicellulose. The complex is permeated with cellulose microfibrils and protein extensin (Fig. 1) [1, 2]. All its components are linked to each other by ester, salt, combined, and hydrogen bonds. Each component possesses valuable physicochemical properties with a good potential for food industry [3–7].
Pectins have the most attractive and numerous functional properties among all the carbohydrates of plant cell walls [5, 8]. They owe these useful properties due to their molecular structure. In their native form, pectins have a water-insoluble supramolecular structure called the protopectin complex. The structure is an extended and highly branched linear and lateral network of polymer fragments (Fig. 2). Lateral branches also have a complex structure and can be interconnected with salt and borate bonds [9–18].
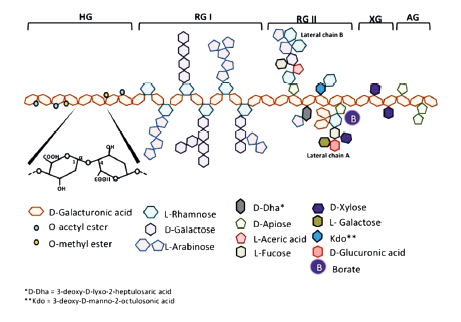
Contemporary science knows eight types of fragments of the protopectin complex: homo-galacturonan, rhamnogalacturonan-I, rhamnogalacturonan- II, xylogalacturonan, apiogalacturonan, and arabinogalacturonan [19].
Homogalacturonans are linear polymeric fragments of α-D(+)-galacturonic acid residues, linked by (1 → 4)-glycosidic bonds (Fig. 3) [19, 20]. Each residue contains a carboxyl group, which naturally may exist in a free, esterified, or amidated state. Free carboxyl groups are capable of dissociation, while acquiring a partial negative charge. Carboxyl groups esterified with methanol demonstrate inactivated charge formation. Amidated carboxyl groups, due to the donor-acceptor bond of the lone-pair electrons, accept cation H+ and acquire a partial positive charge.
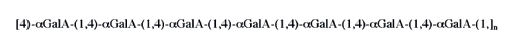
In positions C1 and C2, hydroxyl groups can form glycosidic bonds with the residues of xylose, ribose, arabinose, and galactose, as well as ester bonds with carboxylic acids and aromatic compounds. The state and total amount of carboxyl groups in the pectin molecule fragment define the physicochemical properties of pectins, while the degree and the nature of the substitution of hydroxyl groups define the inhibition degree.
The practical use of pectins depends on the chemical structure of homogalacturonans.
Ramnogalacturonan-I is the second most common fragment of pectins. Its content can reach 45% in sugar beet pectin [5, 19, 20]. These fragments include sequences from the residue of α-L-rhamnose and α-D(+)-galacturonic acid, linked by a (1 → 4)-glycosidic bond. In the rhamnosyl residue, the pair can be linked with other pairs or with the end of the homogalacturonan by a rhamnosyl-uronic (1 → 2)-glycosidic bond. In the uronic residue, the pair can be linked with other pair by a rhamnosyl-uronic (1 → 2)-glycosidic bond or with the end of homogalacturonan by a uronic-uronic (1 → 4)-glycosidic bond. As a result, rhamnosyl residues of rhamnogalacturonan-I are the branching zones of the pectin molecule, where free functional groups can form glycosidic bonds with either residues of neutral sugars, or their polymer sequences, i.e. arabinans, galactans, arabinogalactans, and galactoarabinans-I and II (Fig. 4).
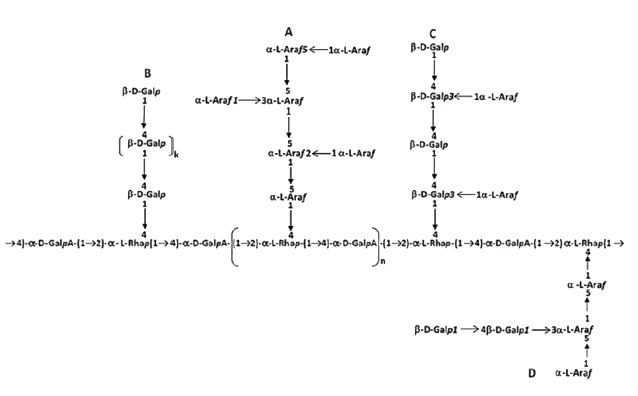
The basis of the protopectin complex of plant tissue cell walls is a network of regions formed by linear sequences of homogalacturonans and rhamnogalacturonans-I. Of course, this assumption excludes two types of lateral branches: the rhamnosilfree lateral branches (rhamnogalacturonan-II), which may contain residues of L-rhamnose and/or α-D(+)-galacturonic acid with proportion of ≤ 2–3%, and branches formed by neutral sugars and their oligo- and polymers [16, 19]. Molecular properties of homogalacturonan fragments define the physicochemical properties of plant pectin. Therefore, enzymatic fragmentation is the most effective method for the protopectin complex. It is a selective hydrolytic cleavage of rhamnosyl-uronide (1 → 2) and (1 → 4) glycosidic bonds.
However, the physicochemical properties of pectin also depend on the polymerization degree of the fragmentation products [21]. The maximal possible degree of polymerization depends on the polymerization degree of the native homogalacturonan fragments in the protopectin complex. In each specific case, the experimental determination of this indicator is a difficult resource- and time-consuming task.
Therefore, a criteria assessment would be the optimal approach to evaluate the potential efficiency of the directed enzymatic fragmentation of a particular plant protopectin complex. Such assessment can also define the boundary conditions that determine the degree of the targeted physicochemical properties of the fermentolysis products. This approach could also determine the conditions for processing any plant tissue or its derivatives. The approach consists of some stepby- step stages. The first stage was a system of criteria for assessing the transformation potential of a plant biopolymer complex [22].
As a next stage, the present research objective was to
develop a system of criteria for assessing the enzymatic
transformation potential of a plant biopolymer complex
as in the case of pectin substances. The research
included the following tasks:
– developing the abovementioned assessment criteria
system, based on the use of zoned criteria space;
– developing a system of boundary conditions for the
classification of plant raw materials according to the
applicability of the enzymatic transformation of its
protopectin complex.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
The protopectin complex of the plant tissue consists of three main types of fragments: homogalacturonan, rhamnogalacturonan-I, and rhamnogalacturonan-II. The latter type was disregarded as its mass fraction in the protopectin complex is ≤ 2%.
Rhamnogaracturonan-I has linear polynalacturonan sites. As a result, the homogalanic component of the protopectin complex can be considered as part of rhamnogalacturonan-I fragments.
A pectin molecule can be classified as rhamnogalacturonan-I only if, in addition to the homogalacturonan component, it contains at least one branch formed by at least one rhamnosyl residue. Consequently, a polymer molecule has at least two homogalacturonan regions with at least one terminal link (rhamnosyl residue) each.
Linear and homogalacturonan regions of the molecular network alternate in the protopectin complex in a particular order. This order presumably depends on the taxonomy of the raw material and the function of the plant parts. The structural features of the fragments of rhamnogalacturonan-I are such that the natural boundaries of the homogalacturonan regions are L-rhamnose residues connected to the terminal uronid links (1 → 2) and (1 → 4) by glycosidic bonds. The fragment can be roughly described by the following sequence: “terminal link of homogalacturonan – rhamnose residue (the branching starts) – branching site – rhamnose residue (the branching ends) – homogalacturonan region – … – section of homogalacturonan – rhamnose residue (the branching starts) – branching site – rhamnose residue (the branching ends) – terminal link of homogalacturonan”.
In the simplest case, the rhamnogalacturonan-I fragment has only one branching site (br = 1). Depending on its structure, the rhamnogalacturonan-I can include only one rhamnosyl residue (zRh = 1). In a more complex case, the rhamnogalacturonan-I may contain several rhamnosyl residues (zRh = q, where q = 1, 2, 3, ...), which alternate with galacturonid residues (Fig. 5).
The number of branching sites may also depend, to some extent, on the plant species and the functional type of the plant tissue.
Figure 5 features no fragments of rhamnogalacturonan- I as their lateral branches are represented mainly by the nonuronic component.
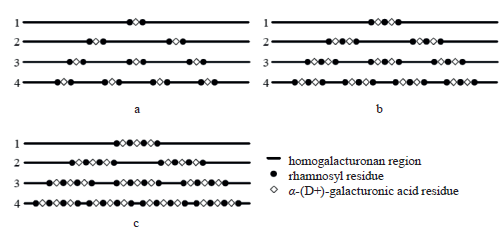
The conditional assumption is that the uronidecontaining part of rhamnogalacturonan-I is completely determined by the following variables: nHG is total homogalacturonan sites, nRh is total rhamnosyl units in the branching sites, nGalA(b) is total uronid residues in the branching sites, zRh is number of rhamnosyl residues per branching site, zGalA(b) is number of uronid residues per branching site, and nbr is total branch sites. Table 1 demonstrates the numerical values of the variables in particular cases of the distribution of homogalacturonan and branching sites in Fig. 5.
The ratios in Table 1 can be expressed by the following formulae:
![]()
![]()
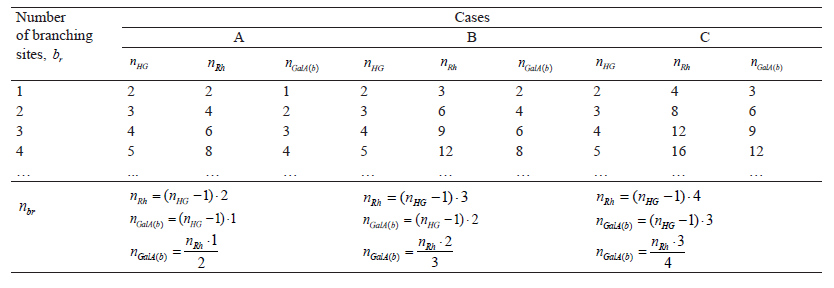
The structure of the rhamnogalacturonan-I fragments suggests that the main structural unit is the amount of rhamnosyl residues in the branching sites. As a result, formulae (1) and (2) take the following form:
![]()
![]()
![]()
Thus, the final formula (4) is:
![]()
These dependences give an approximate quantitative idea of the structure of rhamnogalacturonan-I. For their practical use, they have to be linked to the real chemical composition of a particular raw material.
The line of reasoning follows the next path.
Considering that the molecular weight of the rhamnosyl residue is MRh (Da) and the mass fraction of rhamnose in the composition of the natively insoluble part of the raw material is ωRh (%), the amount of rhamnosyl residues in the mass of the natively insoluble part of the raw material m (g) can be calculated according to the formula below:
![]()
where a is the atomic mass unit (1.66053892×10–24 g/Da). A combination of formulae (6) and (7) gives the number of moles of α-D(+)-galacturonic acid residues in the branch sites:
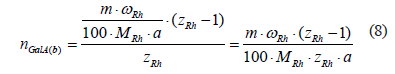
Consequently, the mass fraction of α-D(+)-galacturonic acid residues in the insoluble part of the raw material in the branching sites is:
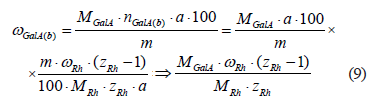
where MGalA is the molar mass of α-D(+)-galacturonic acid residue, Da.
The conditional assumption is that all the residues of α-D(+)-galacturonic acid in the insoluble part belong exclusively to the protopectin complex and are present only in the composition of homogalacturonan fragments and branch points of rhamnogalacturonan-I. Then, the mass fraction of α-D(+)-galacturonic acid residues in homogalacturonan fragments can be calculated as follows:
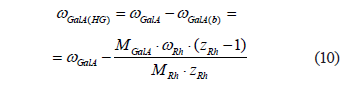
As the plant tissue grows, the protopectin complex of cell walls and intercellular spaces changes continuously. As a result, the structure of the complex becomes heterogeneous. Assuming that all homogalacturonan regions of the protopectin complex are a native component of rhamnogalacturonan fragments, the whole protopectin complex can be represented as consisting almost entirely of rhamnogalacturonan-I fragments. The length of the homogalacturonan regions differs in different parts of the protopectin complex. Consequently, a particular homogalacturonan molecular mass is in fact a certain mean value. The molecular weight of any arbitrarily taken (i-th) homogalacturonan region of the protopectin complex is related to its polymerization degree by the following ratio:
![]()
where ki is the polymerization degree of the i-th homogalacturonan region.
Consequently, the formula for the average molecular weight of homogalacturonan sites is as follows:
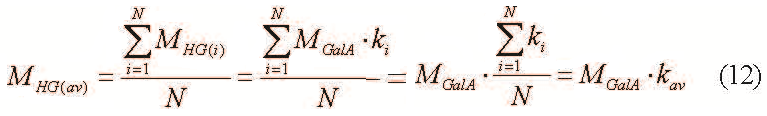
where kav – average polymerization degree of homogalacturonan regions and N – total homogalacturonan regions amount.
The mass fraction of the homogalacturonan component in the insoluble part can be expressed as follows:
![]()
A combination of formulae (3) and (13) gives the following result:
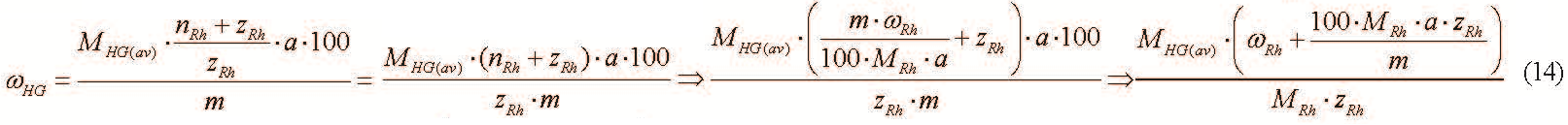
However, the following inequation occurs at m ≥ 10−6g and zRh ≤ 103:
![]()
which makes it possible to disregard the sum of ![]() as insignificant, in which case formula (14)
can be simplified as follows:
as insignificant, in which case formula (14)
can be simplified as follows:
![]()
The mass fraction of homogalacturonan fragments and the mass fraction of α-D(+)-galacturonic acid residues that make up the homogalacturonan fragments are the same, which leads to the following identical equation:
![]()
Added to formula (12), the equation assumes the following form:
![]()
Applying formula (17) to kav makes it possible to calculate the average polymerization degree of homogalacturonan regions in the protopectin complex:
![]()
Thus, the mass fractions of galacturonide and rhamnosyl residues in the plant cell can help to determine the average polymerization degree of the homogalacturonan regions in the protopectin complex.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Let the dimentionless criterion ν is uniquely determined on the basis of chemical analysis of the native water-insoluble plant tissue component:
![]()
As a result, formula (18) looks as follows:
![]()
In (20), constituent MRh/MGalA is constant. Subsequently, formula (20) is a mathematical description of functional dependence kav = f (v, zRh ) (Fig. 6). Thus, analytically obtained ωRh and ωGalA can define the weighted average degree of polymerization of homogalacturonan regions of pectins.
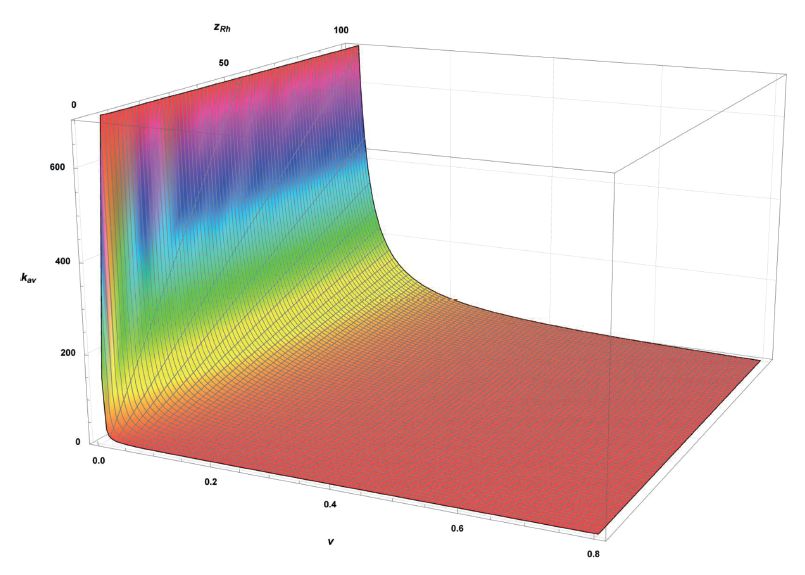
In a same time, homogalacturonan regions in the rhamnogalacturonan fraction of pectin are possible only at kav ≥ 1.
As a result, the rational condition for criterion ν is:
![]()
Provided that there are homogalacturonan regions in
the rhamnogalacturonan fraction of pectin substances,
the range for determining the numerical values of this
criterion can be represented as ![]() . The functional dependence can be reduced to a criterion space in
coordinates ν and zRh, where kav is boundary zoning conditions (Fig. 7).
. The functional dependence can be reduced to a criterion space in
coordinates ν and zRh, where kav is boundary zoning conditions (Fig. 7).
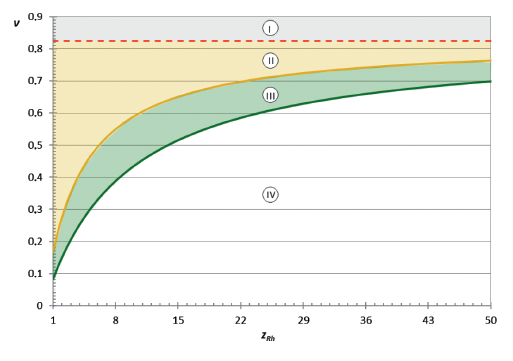
Within this criterion space, zone I is the absence of homogalacturonan regions in pectins. Zone II is the presence of regions with the weighted average polymerization degree of homogalacturonan region in the range of 1–5; zone III – 5–10; and zone IV – ≥ 10.
Homogalacturonan regions with kav > 10 are of their own practical importance. Therefore, the use of homoenzyme preparations for fragmentation of the native protopectin complex makes sense only for plant tissues in zone IV. In other cases, the use of homogalacturonan-specific enzyme preparations for protopectin complex fragmentation has no sense.
The new criteria-based approach makes it possible to unambiguously define the effectiveness of targeted enzymatic fragmentation of the plant protopectin complex within the boundary conditions that determine the degree of the targeted physicochemical properties of the final product. This approach is universal and represents the second stage of the decision tree started in [22] as a science-based technology for pectin production.
ВЫВОДЫ
The research produced a criteria space to assess the potential effectiveness of the homoenzymatic transformation of a plant biopolymer complex as in the case of pectin substances. The method was based on a two-dimensional criteria space, zoned according to the key factor, i.e. the targeted polymerization degree of homogalacturonan fragments in the native protopectin complex.
We found that the compliance with the first criteria zone (at kav ≥ 10) determined the feasibility of using homogalacturonan-specific enzyme preparations to isolate of homogalacturonan (targeted) regions of the plant protopectin complex. The compliance with the second criteria zone (at 1 ≤ kav < 10) determined the expediency of non-enzymatic fragmentation of the protopectin complex. The compliance with the third zone (at kav < 1) meant that the fragmentation of the protopectin complex would neither increase the mass fraction of pectin substances in the medium, nor release pectins. The new criteria approach is an integral part of the technologies for obtaining pectin and its products with targeted physical and chemical properties.
КОНФЛИКТ ИНТЕРЕСОВ
The authors state that there is no conflict of interests related to the publication of this article.ФИНАНСИРОВАНИЕ
The research was part of state assignment of the V.M. Gorbatov Federal Research Center for Food Systems of RAS (Russian Academy of Sciences).СПИСОК ЛИТЕРАТУРЫ
- O’Neill MA, York WS. The composition and structure of plant primary cell walls. In: Rose JKC, editor. The plant cell wall. Annual plant reviews. Volume 8. Oxford: CRC Press; 2003. pp. 1–54.
- Tian L. Influence of pectin supplementation on feed fermentation characteristics in rats and pigs. Dr. sci. diss. Wageningen: Wageningen University; 2016. 144 p. https://doi.org/10.18174/370098.
- Moreno FJ, Sanz ML. Food oligosaccharides: production, analysis and bioactivity. Wiley-Blackwell; 2014. 552 p.
- Ramawat KG, Mérillon J-M. Polysaccharides: bioactivity and biotechnology. Cham: Springer; 2015. 2241 p. https://doi.org/10.1007/978-3-319-16298-0.
- Donchenko LV, Firsov GG. Pektin: osnovnye svoystva, proizvodstvo i primenenie [Pectin: main properties, production, and application]. Moscow: DeLi print; 2007. 275 p. (In Russ.).
- Lisovitskaya EP, Patieva SV, Tymoshenko NV, Patieva AM. Evaluation of analytical characteristics of the different types of pectin in canned technologies for preventive human nutrition. Vsyo o myase. 2016;(3):32–35. (In Russ.).
- Galstyan AG, Aksyonova LM, Lisitsyn AB, Oganesyants LA, Petrov AN. Modern approaches to storage and effective processing of agricultural products for obtaining high quality food products. Herald of the Russian Academy of Sciences. 2019;89(2):211–213. https://doi.org/10.1134/S1019331619020059.
- Bush PL. Pectin: Chemical properties, uses and health benefits. Nova Science; 2014. 268 p.
- May CD. Industrial pectins: Sources, production and applications. Carbohydrate Polymers. 1990;12(1):79–99. https://doi.org/10.1016/0144-8617(90)90105-2.
- Thakur BR, Singh RK, Handa AK. Chemistry and uses of pectin – A review. Critical Reviews in Food Science and Nutrition. 1997;37(1):47–73. https://doi.org/10.1080/10408399709527767.
- Caffall KH, Mohnen D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydrate Research. 2009;344(14):1879–1900. https://doi.org/10.1016/j.carres.2009.05.021.
- Ovodov YuS. Current views on pectin substances. Russian Journal of Bioorganic Chemistry. 2009;35(3):269–284. https://doi.org/10.1134/S1068162009030017.
- Harholt J, Suttangkakul A, Scheller HV. Biosynthesis of pectin. Plant Physiology. 2010;153(2):384–395. https://doi.org/10.1104/pp.110.156588.
- Sato MF, Rigoni DC, Canteri MHG, Petkowicz CLO, Nosueira A, Wosiacki G. Chemical and instrumental characterization of pectin from dried pomace of eleven apple cultivars. Acta Scientarium – Agronomy. 2011;33(3):383–389. https://doi.org/10.4025/actasciagron.v33i3.7125.
- Srivastava P, Malvia R. Sources of pectin, extraction and its application in pharmaceutical industry – An overview. Indian Journal of Natural Products and Resources. 2011;2(1):10–18.
- Leclere L, Cutsem PV, Michiels C. Anti-cancer activities of pH- or heat-modified pectin. Frontiers in Pharmacology. 2013;4. https://doi.org/10.3389/fphar.2013.00128.
- Muller-Maatsch J, Bencivenni M, Caligiani A, Tedeschi T, Bruggeman G, Bosch M, et al. Pectin content and composition from different food waste streams. Food Chemistry. 2016;201:37–45. https://doi.org/10.1016/j.foodchem.2016.01.012.
- Lara-Espinoza C, Carvajal-Millán E, Balandrán-Quintana R, López-Franco Y, Rascón-Chu A. Pectin and pectin-based composite materials: Beyond food texture. Molecules. 2018;23(4). https://doi.org/10.3390/molecules23040942.
- Yapo BM, Gnakri D. Pectic polysaccharides and their functional properties. In: Ramawat KG, Mérillon J-M, editors. Polysaccharides: bioactivity and biotechnology. Cham: Springer; 2015. pp. 1729–1749. https://doi.org/10.1007/978-3-319-16298-0_62.
- Mohnen D. Pectin structure and biosynthesis. Current Opinion in Plant Biology. 2008;11(3):266–277. https://doi.org/10.1016/j.pbi.2008.03.006.
- Kondratenko VV, Kondratenko TYu. Influence of molecular weight on sorption properties display of pectin substances. New Technologies. 2011;(2):20–26. (In Russ.).
- Kondratenko VV, Kondratenko TYu, Petrov AN, Belozerov GA. Assessing protopectin transformation potential of plant tissue using a zoned criterion space. Foods and Raw Materials. 2020;8(2):349–361. http://doi.org/10.21603/2308-4057-2020-2-348-361.
