

Аннотация
Introduction. Brassica L. vegetables are rich in fiber, minerals, and bioactive compounds. Lactic fermentation can improve their nutritional value. The goal of this study was to evaluate phytase, calcium, phytic acid, total phenolic content, and antioxidants during spontaneous fermentation of white cabbage, red cabbage, and Chinese cabbage.Study objects and methods. The research featured samples of water extract, methanol extract, and brine. The procedure involved monitoring lactic bacteria and pH during cabbage fermentation. Diphenyl-1-picrylhydrazyl radical (DPPH) scavenging assay and cupric reducing antioxidant capacity (CUPRAC) assay were used to measure the antioxidant activity and Folin-Ciocalteau method to determine total phenolic content in the water and methanol extracts. In the brine samples, we studied calcium, phytic acid, and phytase activity.
Results and discussion. The samples of white and red cabbage displayed the highest phytase activity on days 5–10 and had a maximal decrease of phytic acid and increase of calcium concentration, while in Chinese cabbage these processes occurred gradually throughout the fermentation. The total phenolic content in the brine and extracts was very similar for all the cultivars throughout the fermentation process. A continuous release from the solid phase to brine could be observed during the first ten days of fermentation. DPPH and CUPRAC assays revealed a similar phenomenon for the total phenolic content. The antioxidant capacity decreased in the water and methanol extracts and increased in the brine. At the end of fermentation, the red cabbage samples demonstrated a significant increase in the total phenolic content and total antioxidant activity, which was less prominent in the Chinese cabbage. The samples of white cabbage, on the contrary, showed a decrease in these parameters.
Conclusion. Fermentation made it possible to increase the concentration of free calcium in white, red, and Chinese cabbages, as well as improve the antioxidant capacity of red and Chinese cabbages.
Ключевые слова
Phytase activity, total phenolics, antioxidant activity, lactic acid bacteria, fermentation, cabbageВВЕДЕНИЕ
Brassicaceae family comprises a large number of plant species distributed all over the world except Antarctica. This family includes approximately 338 genera and 3700 species [1]. Fresh or preserved, cabbage has been part of human diet since ancient times [2].
Brassica oleracea L. and Brassica rapa L. are the most popular representatives of Brassica vegetables. They are almost completely edible, e.g. leaves, inflorescence, root, stem, and seed. Their excellent adaptability makes it possible to cultivate them in different seasons and environments. In the Occident, consumers prefer B. oleracea var. capitata, especially white cabbage and red cabbage. Oriental cuisine features mostly B. rapa var. glabra Regel (Chinese cabbage) or B. rapa ssp. chinensis (L.) Hanelt (pak choi) [3].
Brassica vegetables have low concentrations of protein and fat, which makes them popular low calorie foods. In addition, they are rich in fiber, such minerals as calcium and iron, and such bioactive compounds as polyphenols and glucosinolates [4]. Calcium is essential for human diet. Dairy products are great sources of calcium, both qualitatively and quantitatively. However, people with lactose intolerance and vegans refrain from dairy products, which makes Brassica vegetables an excellent source of the recommended daily calcium intake. Indeed, cabbage has high concentrations of calcium, iron, selenium, copper, manganese, and zinc. Unfortunately, it also contains phytates that may form complexes with calcium, thus reducing its bioavailability and nutritional value [5].
The past decade has seen an increase in scientific interest to the antioxidant properties of dietary plant polyphenols. These secondary metabolites can act as reducing agents (free radical terminators), metal chelators, singlet oxygen quenchers, and hydrogen donors [6]. Furthermore, epidemiological studies strongly suggest that long term consumption of plant polyphenols prevents degenerative diseases associated with oxidative stress [7]. Some recent studies also showed that cruciferous vegetables decrease the risk of several types of cancer, which makes cabbage a functional food [2].
Cabbage can be consumed raw as part of salads, condiments, or juice. It can be subjected to thermal processing, e.g. steaming, boiling, roasting, microwaving, etc., or fermentation (sauerkraut, kimchi, etc.) [2]. Recent studies demonstrated that Brassica vegetables lose their nutrient and health-promoting properties if overheated during domestic cooking [4, 8]. However, fermentation is known to enhance their nutritional properties [9]. Fermentation is one of the oldest ways of food processing and preservation. It is a spontaneous process carried out by lactic bacteria present in vegetables tissues. Fermentation increases the safety, sensory properties, and shelf-life of foods. It also promotes the release of bioactive compounds and reduces anti-nutritional factors [10].
Food safety and shelf-life are associated with microbial competition and the synthesis of inhibitory metabolites, such as lactic acid, acetic acid, hydrogen peroxide, diacetyl, ethanol, bacteriocins, and biosurfactants [10]. Lactic fermentation improves the nutritional value of cabbage, as well as its antioxidant activities. Lactic fermentation reduces phytates, thus improving the bioavailability of essential dietary nutrients, such as minerals, e.g. Ca2+, Zn2+, Mg2+, Mn2+, and Fe2+/3+, proteins, and amino acids [11].
Different databases feature the same nutritional data on Brassica vegetables (energy, fat, protein, mineral content, and carbohydrates) [12]. However, the situation is very different when it comes to the content of bioactive compounds [2]. The profile and concentration of phytochemicals depend on the cultivar, fertilization, agricultural conditions, environment, sowing season, and processing [13]. Furthermore, different studies report different effects of fermentation on the total phenolic compound and antioxidant activity [4, 9, 10, 14]. So far, no studies have featured the changes in the total phenolics and antioxidant activity that occur between cabbage tissue and brine.
The research objective was to evaluate the effect of spontaneous fermentation on: 1) phytase activity, calcium, and phytic acid concentrations; 2) total phenolic content and antioxidants activity of methanol extracts, water extracts, and brine throughout the fermentation of three Brassicaceae cabbages harvested in Patagonia (Argentina).
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Preparing the ferments. Chinese cabbage (Brassica rapa var. glabra Regel), white cabbage (Brassica oleracea var. capitate f. alba), and red cabbage (B. oleracea var. capitata f. rubra) were obtained from a local farm of Valle Inferior del Río Chubut located in Patagonia, Argentina. The cabbages were planted in March 2020 and harvested in June 2020. Before the fermentation, each cabbage head was stripped of dry outer leaves. The cleaned cabbage heads were chopped in a shredder into 2 mm thick strips and mixed with 3.0 % (w/w) of salt. Sterile water homogenized the medium (5 mL/100 g of cabbage). Each cabbage was spontaneously fermented at 18°C for 30 days. The fermentation was performed in duplicate.
Fermentation parameters. The total content of lactic bacteria and pH were monitored during the fermentation process on days 0, 1, 2, 3, 4, 5, 10, 15, 20, 25, and 30. At the beginning of the process, these parameters were examined after 6 and 12 h. The pH of the ferments was measured using a pH meter (model Orion 410A). The lactic bacteria count was monitored by incubating on MRS agar at 30°C for 48 h [15]. The results were expressed as colony forming units per milliliter of experimental sample (CFU/mL).
Preparing the solvent extracts and brine. During fermentations, the solid and liquid samples were withdrawn on days 0, 1, 3, 5, 10, 15, 20, 25, and 30. To prepare the solvent extracts, solid samples were dried at 37°C until constant weight to avoid degradation of thermal-sensitive compounds. After that, they were ground. Methanol and distilled water (1:10 m/V dilution) were used to prepare the extracts. For the methanol extract, the mixes were incubated for 3 h at 37°C under stirring. For the water extract, they were autoclaved for 15 min at 120°C. Both extracts were centrifuged at 13 000×g for 15 min at 25°C. The supernatants were stored at –20°C, while the brine samples (liquid material) were stored at –20°C.
Measuring calcium. The o-cresolftaleín complexone colorimetric method was used to determine the amount of calcium in the cabbage brines. Briefly, 50 μL of sample were mixed with 950 µL of reaction mix composed by 3.7 mM of cresolphtalein complexone and 0.2 mM of amino methyl propanol solution (pH 11). The calcium content in the brines was determined against the calcium standard curve (0–55 µg Ca/mL). The absorbance was measured at 570 nm using a Jenway spectrophotometer (UK). The results were expressed as mg calcium per 100 mL brine (mg Ca/100 mL).
Phytic acid determination. The content of phytic acid was evaluated using an enzymatic method kit (Megazyme International, Ireland), based on its hydrolysis and further determination of free phosphorus. The procedure followed the manufacturer’s instructions. The phosphate released from phytic acid was measured using a modified colorimetric molybdenum blue assay described by McKie et al. [11]. The color reagent was prepared with a solution of 0.6 M sulfuric acid (32 mL/L), ammonium molybdate (5 g/L), and ascorbic acid (20 g/L). After enzymatic treatment, 1.0 mL of color reagent was added to 50 µL of supernatant. The system was incubated for 30 min at 50°C, and the absorbance was measured at 820 nm. A standard curve was constructed with dipotassium phosphate (K2HPO4) (0–0.4 µg/mL). The results were expressed as mg K2HPO4/100 mL brine. The concentration of phytic acid was calculated on the basis of free phosphorus using the formula suggested by McKie et al. [11].
Phytase activity of the brine. Phytase activity was determined by measuring the amount of inorganic phosphate released from sodium phytate as proposed by De Angelisa et al. [16]. Briefly, 180 µL of reactive contained 5 mM of sodium phytate and 200 mM of sodium acetate buffer (pH 5.0). This amount was added to 20 µL of brine. After 15 min of incubation at 37°C, the reaction was stopped by adding an equal volume of 15% trichloroacetic acid. Afterward, the phosphate released was determined by the previously described ammonium molybdate method. One unit of phytase activity was defined as 1 µmol of phosphate produced per min per mL of brine under the assay conditions. The results were expressed as milli-units (mU).
Measuring the total phenolics. The total phenolic content was determined using the Folin-Ciocalteau reagent according to previously published procedures, with minor modifications [17]. An aliquot of 50 μL of extract was mixed with 100 μL of Folin-Ciocalteu´s phenol reagent and kept for 10 min. Then, Na2CO3 (1.0% m/V; 1.0 mL) was added and kept for 90 min at 25°C. The absorbance was measured at 750 nm. A calibration curve was based on gallic acid as standard. The results were expressed as milligram gallic acid equivalents per 100 g of dry weight (mg GAE/100 g DW).
Determination the antioxidant activity. Diphenyl1-picrylhydrazyl (DPPH) radical scavenging assay. The free radical scavenging activity of the samples was evaluated by 1, 1-diphenyl-2-picryl-hydrazyl (DPPH) method as described by Chen et al., with some modifications [18]. Briefly, 900 µL of an ethanolic DPPH solution (100 μM) was added to 100 µL of sample at various concentrations. After 30 min of incubation in the dark at 25°C, the absorbance was measured at 517 nm using a spectrophotometer. A standard curve was constructed with Trolox as a reducing agent (15–250 µg/mL). The results were expressed as milligram Trolox equivalents per 100 g of dry weight (mg TE/100 g DW).
Cupric reducing antioxidant capacity (CUPRAC) assay. Cupric reducing antioxidant power (CUPRAC) was used to determine the antioxidant capacity of the sample as described by Gouda et al., with minor modifications [19]. An aliquot of 100 μL of sample was mixed with 900 µL of reaction mix. The reaction mix consisted of 2 mL of Neocuproine solution (5 mM), 1 mL of Cl2Cu (0.01 M), and 3 mL of acetate buffer (50 mM, pH 5.0). After shaking and incubating for 1 h in the dark, the mix was tested for absorbance at 450 nm. A calibration curve was prepared using Trolox as standard (15–250 µg/mL). The results were expressed as mg of Trolox equivalent per 100 g of dry weight (mg TE/100 g DW).
Total antioxidant capacity. The total antioxidant capacity of the ferments was calculated by adding partial antioxidant activity of extracts and liquid phase (brine) contained in 100 g of edible material to simulate the antioxidant activity per sample. The same procedure was repeated for each vegetable and antioxidant parameter, i.e. DPPH, CUPRAC, and total phenolics. The results were expressed as milligram Trolox equivalents per 100 g of fresh weight ferment (mg TE/100 g FW).
Statistical analysis. All assays were carried out in duplicate, unless mentioned otherwise. The data were analyzed by ANOVA, and the means were compared by the minimum significant difference test at P < 0.05, using the Statgraphics Centurion XVI software.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Fermentation parameters. Lactic bacteria and pH helped monitor the evolution of the fermentation process. Spontaneous fermentation of cabbage relies on autochthonous lactic bacteria present on the raw substrate. Organic acids decrease pH and increase the titratable acidity of the raw material.
The pH of raw white cabbage, red cabbage, and Chinese cabbage were 6.0, 5.9, and 6.1, respectively (Fig. 1). The samples of red and Chinese cabbage demonstrated a similar decrease in pH. In both cultivars, the lowest values were observed on day 4 and remained stable over 30 days (Figs. 1b and 1c). The white cabbage showed no sharp decrease of pH during fermentation. The lowest value was achieved on day 10 and remained stable (Figs. 1a vs 1b and 1c).
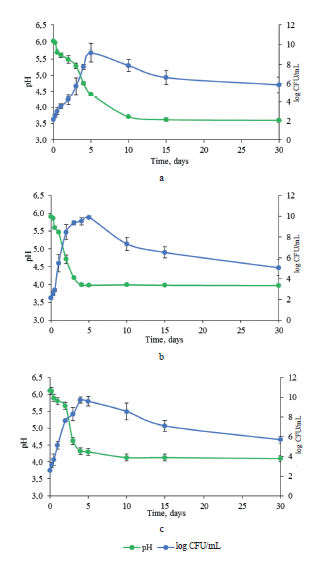
The initial population of lactic bacteria was 2.1, 2.1, and 2.5 log CFU/mL in the white, red, and Chinese cabbages, respectively (Fig. 1). This trend confirms previous reports by R. Di Cagno et al. and J. Beganović et al. [10, 20]. While the highest count was observed on day 5 (9.9 log CFU/mL), the red cabbage sample approached its maximal counts on day 3 (9.4 log CFU/mL) (Fig. 1b). A similar curve was observed for the Chinese cabbage fermentation; however, the maximal counts were detected after day 4 (9.7 log CFU/mL) (Fig. 1c). Regarding the white cabbage, lactic bacteria population increased slower than in other samples and reached its maximum (9.2 log CFU/mL) on day 5 (Fig. 1a). In all the cases, once the peak was reached, the bacteria populations began to decrease. On day 30, the lactic bacteria cell counts were 5.0, 5.9, and 5.7 log CFU/mL for red cabbage, white cabbage, and Chinese cabbage, respectively (Fig. 1).
Calcium, phytic acid, and phytase activity. Figure 2 shows the changes in the phytase activity and calcium and phytic acid concentrations that occurred in the brine during fermentation. The raw samples of red and Chinese cabbage (Figs. 2b and 2c, respectively) contained comparable amounts of water-soluble calcium, whereas the white cabbage appeared to have a much lower concentration (Fig. 2a).
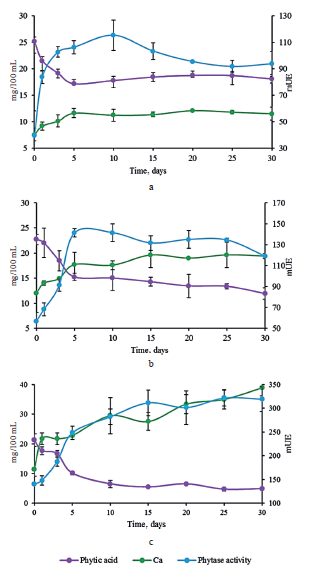
The initial level of phytic acid was almost the same for all three cultivars. The raw sample of Chinese cabbage showed the highest phytase activity (Fig. 2c). The initial specific activities of white and red cabbages were 39.54 ± 18.67 (Fig. 2a) and 56.71 ± 8.20 mU (Fig. 2b), respectively. The enzymatic activity was supplied exclusively by vegetal tissue during early fermentation, and then bacterial phytase brought about phytate hydrolysis [21].
The highest phytase activity was detected between days 5 and 10 in the samples of white and red cabbage, when the population of lactic bacteria reached its maximum (Figs. 2a and 2b). After that, the values remained constant. In the sample of Chinese cabbage, the maximal activity was detected on day 10 (Fig. 2c), which coincided with the maximal viable cell count of lactic bacteria.
As the fermentation process advanced, the phytate concentration decreased and the amount of soluble calcium increased in all the samples. This phenomenon was more pronounced in Chinese cabbage when the phytase activity had its highest value. The lowest phytate concentration and the highest calcium concentration were achieved on day 30. The assays for all the samples proved that the highest phytase activity occurred under acidic conditions.
Phytic acid (myo-inositol-6-phosphate) is the major storage form of phosphorous and represents 50–85% of total phosphorous in plants [21]. This compound and its derivatives are the main inhibitors of divalent mineral absorption in the gastrointestinal tract due to the formation of insoluble and indigestible complexes [22]. Hence, it may decrease the calcium bioavailability in cabbage [21]. However, this point of view is now controversial since several studies demonstrated that the myo-inositol-6-phosphate consumption may be associated with some health benefits. The antinutrient effect of phytic acid has not been fully demonstrated in vivo. On the other hand, phytic acid exerts antiinflammatory and anticancer activities and diminishes the risk of osteoporosis [23].
Phenolic compounds. Phenolic composition and antioxidant activity depend mainly on the type of extraction solvent. The choice of solvent depends mainly on the chemical nature and polarity of the compounds to be extracted. Methanol and water are widely used as solvents in vegetable and plant tissues [14]. In this study, methanol and water helped measure phenolic compounds and antioxidant activity in the cabbage samples during fermentation.
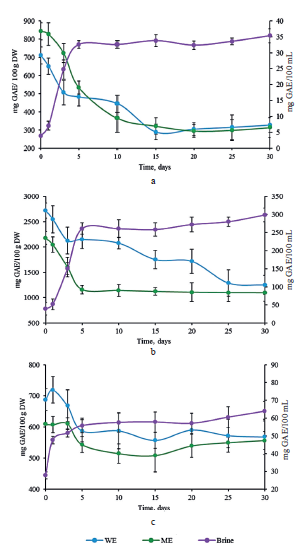
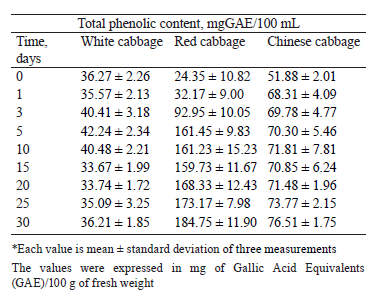
Figure 3 shows the total phenolic content in the extracts (methanol and water) and brines of white, red, and Chinese cabbages. Regarding the white cabbage sample, the water and methanol extracts exhibited a similar total phenolic content. However, the total phenolic content in the water extracts of red and Chinese cabbages was much higher than in the methanolic extract (Figs. 3b and 3c). Probably, the solubility of phenolic compounds depended on extraction conditions, e.g. the chemical structure of solvents, dielectric constant, time, temperature, phytochemical properties, etc. However, thermal treatment is known to damage some phenolics [24].
The total phenolic content in the extracts and brine of red cabbage was higher than in the samples of white and Chinese cabbage. This trend was in agreement with previous studies. For instance, Tabart et al. [25] reported 1851 mg GAE/100 g DW in red cabbage; Vicas et al. [26] – 980–1220 mg GAE/100 g DW in white cabbage; Seong et al. [27] – 347.46 ± 32.17 mg GAE/100 g DW in Chinese cabbage. In vegetables, phenolics exist mostly in conjugated forms through hydroxyl groups with sugar as glycosides. Lactic bacteria possess an enzymatic battery that can convert phenolics to aglycone forms, which are simpler and biologically more active [28]. Furthermore, during fermentation, pectic enzymes may soften cabbage texture, thus releasing phenolics compounds from the solid to the liquid phase [27].
Lactic fermentation promoted a significant decreased in the total phenolic content in the red and white cabbage extracts (methanol and water) after 3–5 days of incubation (Figs. 3a and 3b). Afterward, the total phenolic content dropped slowly and remained almost constant until the end of lactic fermentation. The total phenolic content in the methanol and water extracts decreased almost by half. On the contrary, the brine samples demonstrated a significant increase between days 3 and 5, and then the concentration remained almost stable until the end of storage. The Chinese cabbage sample showed a slight decrease in the total phenolic content in methanol and water extracts throughout the fermentation (Fig. 3c).
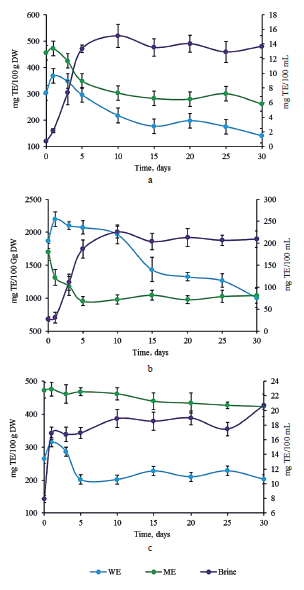
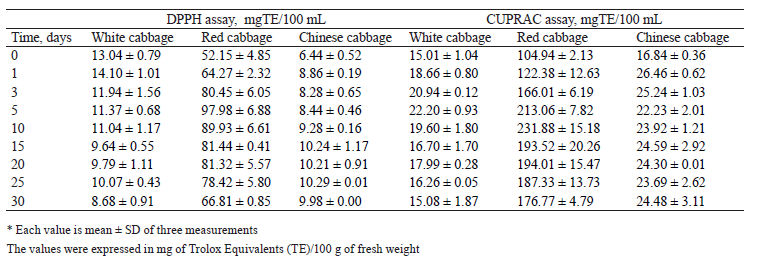
Antioxidant activity. The antioxidant activity was evaluated by DPPH radical scavenging assay and CUPRAC reduction assay. Both are electron transferbased methods, frequently used to determine the antioxidant activities of phenolic compounds [6]. Figure 4 illustrates the antiradical activity of methanol and water extracts against DPPH radical. The methanol extract contained significantly less reduction power than the water extract in all the cabbage samples. Probably, this solvent failed to provide efficient extraction of compounds with antioxidant activity. The raw sample of red cabbage showed the highest antioxidant capacity with 1050.44 ± 71.33 TE/100 g DW and 616.63 ± 49.80 mg TE/100 g DW in water and methanol extracts, respectively (Fig. 4b). Afterwards, these values declined until the end of fermentation.
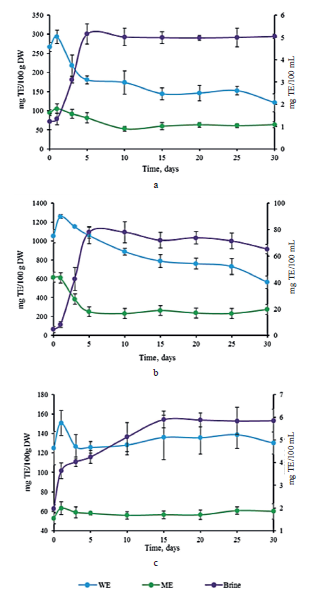
On the contrary, in the brine, the values kept rising until day 5 and then remained stable. The extracts of white and Chinese cabbages displayed a significantly lower DPPH radical scavenging activity than the extracts of red cabbage (Figs. 4a and 4c). These trends confirmed previous reports [29]. The methanol and water extracts of white cabbage exhibited a slight decrease in the antiradical activity, while its brine demonstrated an increase during the first 5 days of fermentation (Fig. 4a). However, no significant differences in the antioxidant activities were observed in the Chinese cabbage extracts. A significant increase was detected in the brine during the first 10 days of fermentation, but it remained constant until the end of fermentation (Fig. 4c).
In all the cases, the values of antioxidant capacity obtained with CUPRAC assay (Fig. 5) were higher than those obtained with DPPH method. This trend could be explained by the ability of CUPRAC method to measure hydrophilic and lipophilic antioxidants simultaneously, while DPPH detects only those molecules that are soluble in organic solvents, particularly in alcohols [30].
The antioxidant capacity of the red and white cabbages decreased significantly in the methanol and water extracts during day 1 and increased significantly in the brine (Figs. 5a and 5b). In the white cabbage, these changes occurred between days 5 and 10. For the red cabbage, the decrease was observed on day 5 in the methanol extract and on day 15 in the water extracts. The maximal value in brine was achieved after 5 days.
Regarding the Chinese cabbage samples, a comparable trend could be observed between the values obtained with DPPH radical scavenging assay and CUPRAC method. The concentration of reducing agents in dry matter decreased slowly in the water extract, while the methanol extract showed no significant differences. A slight but significant increase in the concentration was detected in the brine (Fig. 5c).
The antioxidant capacity presented a sharp increase on day 1 (Figs. 4 and 5). This trend was due to the high driving force produced by concentration gradients of the substance that tends to equilibrate the medium. In this process, water flows from the solid phase to the liquid phase and brings some solutes from the vegetables. This phenomenon is due to transfer rates that increase or decrease until equilibrium is reached [31].
Overall evaluation of total phenolics and antioxidant activity. The total phenolic content and antioxidant activity in the white and red cabbage samples decreased in the dry matter and increased in the liquid phase. This phenomenon was less pronounced in the Chinese cabbage sample. However, these data alone cannot estimate the total variation of the antioxidant capacity throughout the process: both phases contributed to the phenolic content and scavenging activity since the cabbages were not to be consumed dry.
Tables 1 and 2 show the results obtained by adding the values of dry matter and brine. These results can be considered the total polyphenol content and the total antioxidant activity of the fermented cabbages. Regarding the samples of red and Chinese cabbages, the total phenolic content and the total scavenging activity in the water extracts and brine gradually increased and reached plateau after about 5–10 days, which coincided with the highest population of lactic bacteria. In the white cabbage samples, the total phenolic content and the total antioxidant capacity in the water extract and liquid phase exhibited slight changes. By the end of fermentation, the total phenolic content and the antioxidant activity were similar or smaller, in the case of radical scavenging activity measured by DPPH.
To sum up, the fermentation increased the total phenolic content and the antioxidant activity in the liquid phases of red and Chinese cabbages. The red cabbage sample had the highest total phenolic content.
ВЫВОДЫ
Fermentation was able to significantly improve the quality and functionality of Brassica L. cabbages. The test samples showed a significant increase in phytase activity, which promoted the decrease of phytic acid and the increase of free calcium. Fermentation raised the total phenolic content and the antioxidant activity because of the individual contribution of the solid and liquid phases to total scavenging capacity.Вклад авторов
Romina Parada is responsible for conceptualization, methodology, software, validation, formal analysis, investigation, reviewing, proofreading, and visuali- zation. Emilio Marguet is responsible for conceptuali- zation, methodology, formal analysis, investigation, and drafting. Carmen Campos is responsible for conceptualization, software, formal analysis, writingreviewing, and editing. Marisol Vallejo participated in conceptualization, methodology, writing, reviewing, editing, and visualization.КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that there is no conflict of interests regarding the publication of this article.ФИНАНСИРОВАНИЕ
This work received support from the National University of Patagonia San Juan Bosco (UNPSJB) Exchange project codes. National University of Patagonia San Juan Bosco (UNPSJB)(SCT PI Nº1519), Secretariat of University Policies for the National Ministry of Education (UNPSJB7744), Secretariat of University Policies for the National Ministry of Education Please add: National Promotion Agency Scientific and Technological (PICT 2019-01348), University of Buenos Aires (UBA) (20020170100065BA), and the National Research Council of Argentina, CONICET (11220150100042CO).СПИСОК ЛИТЕРАТУРЫ
- Al-Shehbaz IA, Beilstein MA, Kellogg EA. Systematics and phylogeny of the Brassicaceae (Cruciferae): An overview. Plant Systematics and Evolution. 2006;259(2–4):89–120. https://doi.org/10.1007/s00606-006-0415-z.
- Šamec D, Salopek-Sondi B. Cruciferous (Brassicaceae) vegetables. In: Mohammad Nabavi S, Silva AS, editors. Nonvitamin and nonmineral nutritional supplements. Academic Press; 2019. pp. 195–202. https://doi.org/10.1016/b978-0-12-812491-8.00027-8.
- Lee H, Oh I-N, Kim J, Jung D, Cuong NP, Kim Y, et al. Phenolic compound profiles and their seasonal variations in new red-phenotype head-forming Chinese cabbages. LWT – Food Science and Technology. 2018;90:433–439. https://doi.org/10.1016/j.lwt.2017.12.056.
- Kusznierewicz B, Śmiechowska A, Bartoszek A, Namieśnik J. The effect of heating and fermenting on antioxidant properties of white cabbage. Food Chemistry. 2008;108(3):853–861. https://doi.org/10.1016/j.foodchem.2007.11.049.
- Jahangir M, Kim HK, Choi YH, Verpoorte R. Health-affecting compounds in Brassicaceae. Comprehensive Reviews in Food Science and Food Safety. 2009;8(2):31–43. https://doi.org/10.1111/j.1541-4337.2008.00065.x.
- Shahidi F, Zhong Y. Measurement of antioxidant activity. Journal of Functional Foods. 2015;18:757–781. https://doi.org/10.1016/j.jff.2015.01.047.
- Pandey KB, Rizvi SI. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Medicine and Cellular Longevity. 2009;2(5):270–278. https://doi.org/10.4161/oxim.2.5.9498.
- Podsedek A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT – Food Science and Technology. 2007;40(1):1–11. https://doi.org/10.1016/j.lwt.2005.07.023.
- Hur SJ, Lee SY, Kim Y-C, Choi I, Kim G-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chemistry. 2014;160:346–356. https://doi.org/10.1016/j.foodchem.2014.03.112.
- Di Cagno R, Coda R, De Angelis M, Gobbetti M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiology. 2013;33(1):1–10. https:/doi.org/10.1016/j.fm.2012.09.003.
- McKie VA, McCleary BV. A novel and rapid colorimetric method for measuring total phosphorus and phytic acid in foods and animal feeds. Journal AOAC International. 2016;99(3):738–743. https://doi.org/10.5740/jaoacint.16-0029.
- PLANTS database [Internet] [cited 2021 Aug 10]. Available from: https://plants.usda.gov.
- Aires A, Fernandes C, Carvalho R, Bennett RN, Saavedra MJ, Rosa EAS. Seasonal effects on bioactive compounds and antioxidant capacity of six economically important Brassica vegetables. Molecules. 2011;16(8):6816–6832. https://doi.org/10.3390/molecules16086816.
- Sun Y-P, Chou C-C, Yu R-C. Antioxidant activity of lactic-fermented chinese cabbage. Food Chemistry. 2009;115(3):912–917. https://doi.org/10.1016/j.foodchem.2008.12.097.
- Di Cagno R, Minervini G, Rizzello CG, De Angelis M, Gobbetti M. Effect of lactic acid fermentation on antioxidant, texture, color and sensory properties of red and green smoothies. Food Microbiology. 2011;28(5):1062–1071. https://doi.org/10.1016/j.fm.2011.02.011.
- De Angelisa M, Gallo G, Corbo MR, McSweeney PLH, Faccia M, Giovine M, et al. Phytase activity in sourdough lactic acid bacteria: purification and characterization of phytase from Lactobacillus sanfranciscensis CB1. International Journal Food Microbiology. 2003;87(3):259–270. https://doi.org/10.1016/S0168-1605(03)00072-2.
- Agbor GA, Vinson JA, Donnelly PE. Folin-Ciocalteau reagent for polyphenolic assay. International Journal of Food Science, Nutrition and Dietetics. 2014;3(8):147–156. https://doi.org/10.19070/2326-3350-1400028.
- Chen Y-C, Sugiyama Y, Abe N, Kuruto-Nima R, Nozawa R, Hirota A. DPPH radical scavenging compounds from Dou-Chi, a soybean fermented food. Bioscience, Biotechnology, and Biochemistry. 2005;69(5):999–1006. https://doi.org/10.1271/bbb.69.999.
- Gouda AA, Amin AS. Copper(II)-neocuproine reagent for spectrophotometric determination of captopril in pure form and pharmaceutical formulations. Arabian Journal of Chemistry. 2010;3(3):159–165. https://doi.org/10.1016/j.arabjc.2010.04.004.
- Beganović J, Kos B, Leboš Pavunc A, Uroić K, Jokić M, Šušković J. Traditionally produced sauerkraut as source of autochthonous functional starter cultures. Microbiological Research. 2014;169(7–8):623–632. https://doi.org/10.1016/j.micres.2013.09.015.
- Gupta RK, Gangoliya SS, Singh NK. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. Journal of Food Science and Technology. 2013;52(2):676–684. https://doi.org/10.1007/s13197-013-0978-y.
- Lazarte CE, Carlsson N-G, Almgren A, Sandberg A-S, Granfeldt Y. Phytate, zinc, iron and calcium content of common Bolivian food, and implications for mineral bioavailability. Journal of Food Composition and Analysis. 2015;39:111–119. https://doi.org/10.1016/j.jfca.2014.11.015.
- Duong QH, Lapsley KG, Pegg RB. Inositol phosphates: health implications, methods of analysis, and occurrence in plant foods. Journal Food Bioactives. 2018;1(1):41–55. https://doi.org/10.31665/JFB.2018.1126.
- Arfaoui L. Dietary plant polyphenols: Effects of food processing on their content and bioavailability. Molecules. 2021; 26(10). https://doi.org/10.3390/molecules26102959.
- Tabart J, Pincemail J, Kevers C, Defraigne J-O, Dommes J. Processing effects on antioxidant, glucosinolate, and sulforaphane contents in broccoli and red cabbage. European Food Research and Technology. 2018;244(12):2085–2094. https://doi.org/10.1007/s00217-018-3126-0.
- Vicas SI, Teusdea AC, Carbunar M, Socaci SA, Socaciu C. Glucosinolates profile and antioxidant capacity of Romanian Brassica vegetables obtained by organic and conventional agricultural practices. Plant Foods for Human Nutrition. 2013;68(3):313–321. https://doi.org/10.1007/s11130-013-0367-8.
- Seong G-U, Hwang I-W, Chung S-K. Antioxidant capacities and polyphenolics of chinese cabbage (Brassica rapa L. ssp. Pekinensis) leaves. Food Chemistry. 2016;199:612–618. https://doi.org/10.1016/j.foodchem.2015.12.066.
- Lee N-K, Paik H-D. Bioconversion using lactic acid bacteria: Ginsenosides, GABA, and phenolic compounds. Journal of Microbiology and Biotechnology. 2017;27(5):869–877. https://doi.org/10.4014/jmb.1612.12005.
- Upadhyay R, Sehwag S, Singh SP. Antioxidant activity and polyphenol content of Brassica oleracea varieties. International Journal of Vegetable Science. 2016;22(4):353–633. https://doi.org/10.1080/19315260.2015.1048403.
- Apak R, Güçlü K, Demirata B, Özyürek M, Çelik SE, Bektasog˘lu B, et al. Comparative evaluation of various total antioxidant capacity assays applied to phenolic compounds with the CUPRAC assay. Molecules. 2007;12(7):1496–1547. https://doi.org/10.3390/12071496.
- Yadav AK, Singh SV. Osmotic dehydration of fruits and vegetables : a review. Journal of Food Science and Technology. 2014;51(9):1654–1673. https://doi.org/10.1007/s13197-012-0659-2.
