

Аннотация
Introduction. Public healthcare urgently needs new pharmaceuticals – alternative to traditional antibiotics – that pathogens develop no resistance to. Of special interest in this regard are antimicrobial, ribosomally synthesized bacterial peptides or bacteriocins. In this work, we aimed to study the structure and properties of antimicrobial peptides produced by antagonist microorganisms isolated from the natural objects of the Siberian region.Study objects and methods. The study objects were bacteria isolated from the natural sources of Kuzbass. After culturing bacteria, total protein was precipitated from the culture fluid and separated into fractions by gel permeation HPLC. Their amino acid sequences were determined by MALDI-TOF mass spectrometry. The antibacterial (against Bacillus pumilus and Escherichia coli) and fungicidal (against Aspergillus flavus and Aspergillus niger) properties of the peptides were studied by the disk diffusion method.
Results and discussion. Seven peptides with different amino acid sequences were isolated from the culture fluid of bacteria, five of which had no analogues in the PepBank and Uniprot data banks. The peptide with an amino acid sequence of VMCLARKCSQGLIVKAPLM (2061.66 Da) was homologous to the cysteine membrane protein Giardia lamblia P15, and the peptide with an amino acid sequence of AVPSMKLCIQWSPVRASPCVMLGI (2587.21 Da) showed a homology with the Planctomycetes bacterium I41 peptides. We found antibacterial (against gram-positive and gram-negative bacteria) and fungicidal (against Aspergillus) properties in the peptide fractions.
Conclusion. Antimicrobial peptides produced by bacteria isolated from the natural objects of the Siberian region can be used to create pharmaceuticals as an alternative to traditional antibiotics to treat infectious diseases.
Ключевые слова
Antimicrobial peptide, bacteriocin, fungicide, antagonistic properties, antibiotic resistance, amino acid sequence, mass spectrometry, bacteriaВВЕДЕНИЕ
Pathogenic microorganisms resistant to traditional antibiotics are a serious problem of modern healthcare. There is evidence that over 70% of all pathogenic bacteria are resistant to at least one of the most commonly used antibiotics. Therefore, there is an urgent need for new drugs and therapeutic approaches to overcome their resistance [1–5].
Antimicrobial peptides produced by various organisms from bacteria to mammals are an ideal alternative to antibiotics due to their antimicrobial, antiinflammatory, angiogenic, and immunomodulatory properties, as well as low bacterial resistance [6]. However, their use is limited by toxicity and stability in vivo [7].
Antimicrobial peptides act against various types of pathogens, including Gram-positive and Gram-negative bacteria, viruses, and microscopic fungi, through the destruction of the cytoplasmic membrane, intracellular penetration, and immunomodulation [8, 9]. Structurally, antimicrobial peptides are classified into linear cationic amphipathic peptides and macrocyclic peptides [10]. As a rule, antimicrobial peptides are short peptides consisting of 10–50 amino acids [11, 12]. They have common features despite differing in length, amino acid sequences, and conformation [13]. Typical antimicrobial peptides are composed of positively charged residues such as arginine, lysine, and histidine [14]. Cationic peptides with a positive charge ranging from +2 to +11 can interact with the membranes of microbial cells. Besides, a significant part of antimicrobial peptides is hydrophobic, contributing to the formation of amphipathic secondary or quaternary structures [15].
Antimicrobial peptides have several advantages over traditional antibiotics [16]. First of all, they have a broad spectrum of antimicrobial activity, against even multidrug-resistant pathogens [8, 16]. Secondly, antimicrobial peptides are highly active against gramnegative bacteria, which are more serious targets than gram-positive bacteria [17]. Another advantage is a rather low likelihood of drug resistance.
Bacteriocins are antimicrobial, ribosomally synthesized peptides of bacteria with a low molecular weight [18]. Mostly studied are bacteriocins produced by lactobacilli. They can be roughly divided into four categories: lantibiotics (e.g., nisin); non-antibiotic bacteriocins with good activity against Listeria monocytogenes, as well as pediocins, which make up the largest group; thermosensitive macromolecular proteinaceous bacteriocins; and complex bacteriocins with carbohydrates, lipids, and proteins [19–23]. Of all well-studied bacteriocins of lactobacilli, only nisin is produced commercially [24].
Potential sources of bacteria producing bacteriocins are dairy products, cow rumen, feed, as well as natural objects such as soils, plant waste, rhizosphere of plants, bottom sediments of water bodies, etc. [18, 25, 26].
In our previous studies, we isolated 19 microorganisms from the natural sources of Kemerovo Region (Siberian Federal District, Russia), including 10 species of bacteria (Geobacillus, Bacillus, Lactobacillus, Leuconostoc, and Pediococcus) that showed high antimicrobial activity against Escherichia coli, Salmonella enterica, Staphylococcus aureus, Pseudomonas aeruginosa, Bacillus mycoides, Candida albicans, and Penicíllium citrinum [27–29].
In this study, we aimed to examine the structure and properties of antimicrobial peptides produced by antagonist microorganisms isolated from the natural objects in Siberia.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Our study objects were bacteria isolated from the natural sources of Kuzbass (Table 1).

Microorganism cultures. To obtain enrichment cultures of microorganisms, we crushed the samples of soil, bottom sediments, and plant waste under sterile conditions and rubbed their small amounts on Petri dishes with nutrient agar. The Petri dishes were incubated for three days at 26°C. Two nutrient media were used: lactobacilli were cultured on MRS agar; Bacillus and Geobacillus bacteria were cultured on a medium (pH 7.4 ± 0.2) containing 10.0 g/L casein hydrolysate, 2.5 g/L yeast extract, 5.0 g/L glucose, 2.5 g/L potassium hydrogen phosphate, and 12.0 g/L bacteriological agar.
Pure cultures of microorganisms were obtained from enrichment cultures by streaking. Microorganisms were cultivated on the media described above for 24 h: Lactobacillus, Leuconostoc and Pediococcus bacteria at 37°C, and Bacillus and Geobacillus at 30°C.
At the end of cultivation, cell debris was removed from all suspension cultures. The cultures were centrifuged at 3900 rpm in plastic flasks. The resulting supernatant was dried in a Labcocnco Triad freeze dryer (Labcocnco, USA) at a freezer temperature of –80°С, supernatant temperature of –20°С, and 0.05 mbar vacuum.
Protein fractions. To separate protein into individual fractions, the dried biomass was dissolved in 1 mL of 0.25 M phosphate buffer and the total protein was precipitated by adding 2 mL of concentrated ammonium sulfate solution. The resulting protein suspension was separated by centrifugation at 8000 rpm. The protein precipitate was dissolved in 1 mL of 0.025 M Tris buffer solution (pH 4.5). The precipitate was applied to an Enrich 650 10 mm × 300 mm column (Biorad, USA) for a gel permeation high performance liquid chromatography (HPLC) at 280 nm using a direct injection system. Fractionation was performed using an NGC fraction collector (Biorad, USA).
Additionally, each protein fraction was purified on hydrophobic Amberlite XAD X-6 resins by chromatography. A glass column was filled with 10 g of Amberlite XAD-2 resin equilibrated with 10 mL of 20 mM trifluoroacetic acid solution. A protein solution in an acetate buffer was applied to the column and eluted in a methanol gradient from 0 to 15%, with a gradient rise of 5% for every 10 fractions. Fractions containing proteins were determined by taking 50 μL of each fraction and mixing it with a solution of Bradford’s reagent in a 1:1 ratio. The resulting solution was measured on a Biorad SmartSpec Plus Spectrophotometer (USA). Fractions with an optical absorption of 0.06 or more were selected for further drying and identifying the amino acid sequence by the MALDI-TOF method using a MALDI TOF/TOF BRUKER Autoflex Speed mass spectrometer (Bruker Corporation, USA).
Trypsinolysis. Peptides were precipitated by adding an equal volume of methanol/chloroform mixture to an aliquot of a 200 μL fraction. The resulting precipitate was separated by centrifugation at 4000 rpm. The precipitate was dissolved in 100 μL of 6 M urea solution, to which 5 μL of dithiothreitol (DTT) solution was added to keep for 60 min at room temperature. Then, we added 20 μL of iodoacetamide solution and kept the mixture for 60 min at room temperature. After that, we added 20 μL of a DTT solution and kept the mixture again for 60 min at room temperature. After adding 775 μL of MiliQ H2O and 50 μL of trypsin solution, the mixture was stirred by pipetting and kept in a thermostat at 37°C for 12 h. The enzyme was inactivated by adding 10 μL of trifluoroacetic acid. The peptides were purified by chromatography on C18 cartridges. The reaction mixture was applied to a cartridge and eluted with a solution of 0.1% trifluoroacetic acid in a 1:1 H2O/acetonitrile mixture. Analysis and Top-Dawn sequencing were performed on 1 μL of a purified peptide solution.
The antibacterial properties of the peptides against Bacillus pumilus and Escherichia coli were measured by the disk diffusion method. For this, we used suspensions of night cultures grown on a standard liquid nutrient LB medium with a titer of 0.5. The number of microorganisms (titer) in the suspension was determined by optical density at 595 nm. 200 μL of the pathogen culture was dropped onto a 90 mm Petri dish, rubbed with a sterile spatula by the spread plate method, and left to dry for 20 min under a laminar with the lid ajar. Then, 0.5 cm sterile filter disks soaked in the peptide solutions under study and dried at room temperature for 10 min were placed on the Petri dishes in the radial direction. The Petri dishes were left for 30 min at room temperature and then incubated in a thermostat at 37°C for 12 h. Then, we identified a bacterial inhibition zone around the disc and measured its diameter with a vernier caliper. Ampicillin at a concentration of 5 mg/mL was used as a positive control, and a disc soaked in a liquid medium was used as a negative control.
The fungicidal activity of the peptides against the microscopic fungi Aspergillus flavus and Aspergillus niger was measured by the disk diffusion method. The fungi were cultivated for 7 days, with an inoculation density of 6×107 conidia per 1 mL of medium. The results were analyzed with time intervals (3, 9, 12, 24, 48, 72 h, etc.) and by the fungus growth phase (stationary, accelerated growth, logarithmic), i.e., during the periods of exponential cell growth, decreased growth, and death or autolysis. At the end of the incubation, the inhibition zone around the disc was measured with a vernier caliper (mm), which indicated the degree of biocidal activity or its absence. A negative control was the samples with filters impregnated with the medium, and a positive control was the pharmaceutical preparation Irunin® (Veropharm, Russia) with itraconazole as an active ingredient.
Statistical data were analyzed in Microsoft Office Excel 2007. All the experiments were carried out in triplicate. Statistical analysis was performed using a one-sample Student’s t-test. The differences were considered statistically significant at P < 0.05.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Several protein fractions were isolated from the culture fluid of all the studied samples (Table 2).

According to Table 2, one protein fraction was isolated from the culture fluid of Bacillus subtilis, Lactobacillus plantarum, Lactobacillus casei, and Bacillus caldotenax; two protein fractions from Leuconostoc mesenteroides, Pediococcus acidilactici, and Geobacillus stearothermophilus; three protein fractions from Lactobacillus fermentum; four protein fractions from Pediococcus damnosus; and eight protein fractions from the Pediococcus pentosaceus culture fluid.
The results of the MALDI TOF mass spectrometry of protein fractions are presented in Figs. 1–7. We found some identical mass spectra of protein fractions synthesized by different bacteria.
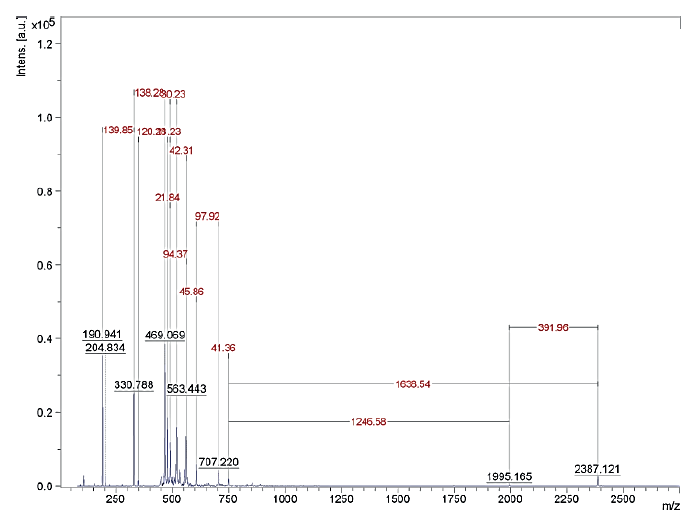

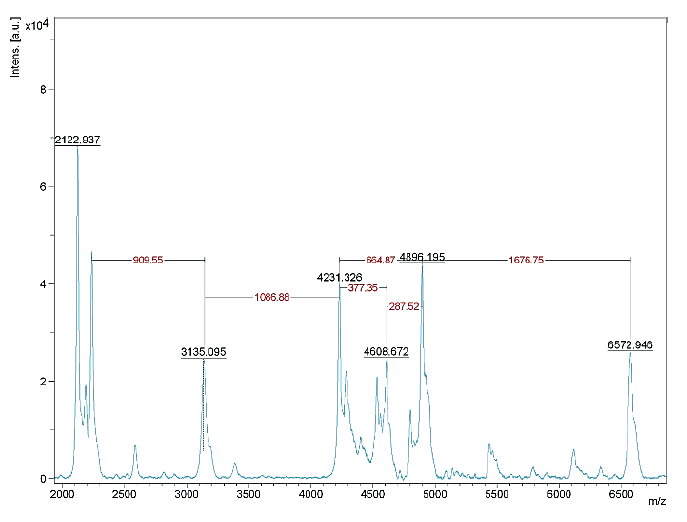
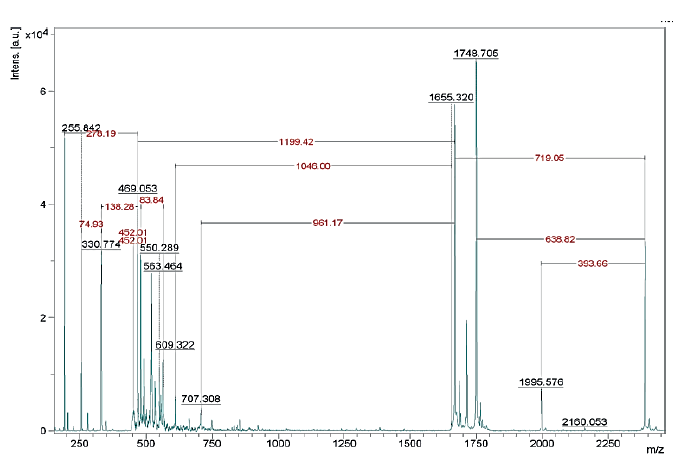
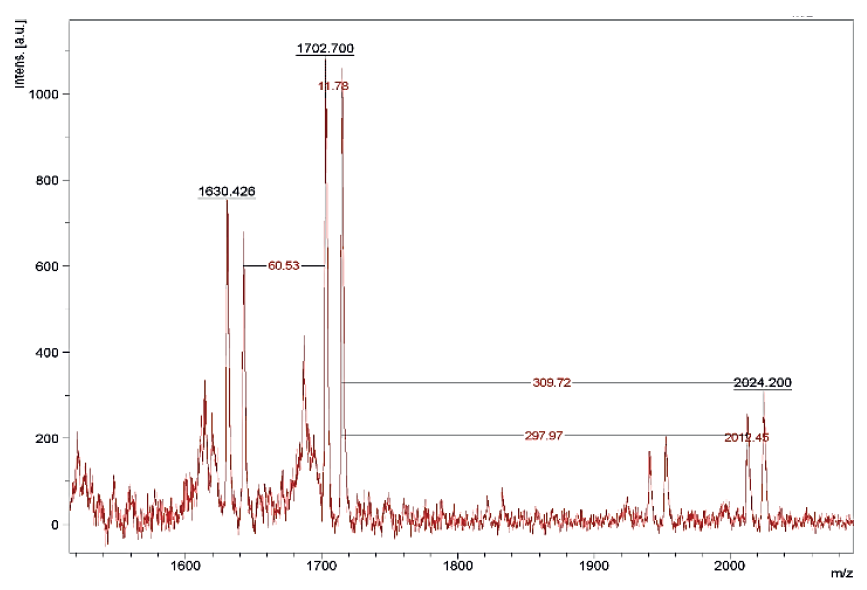
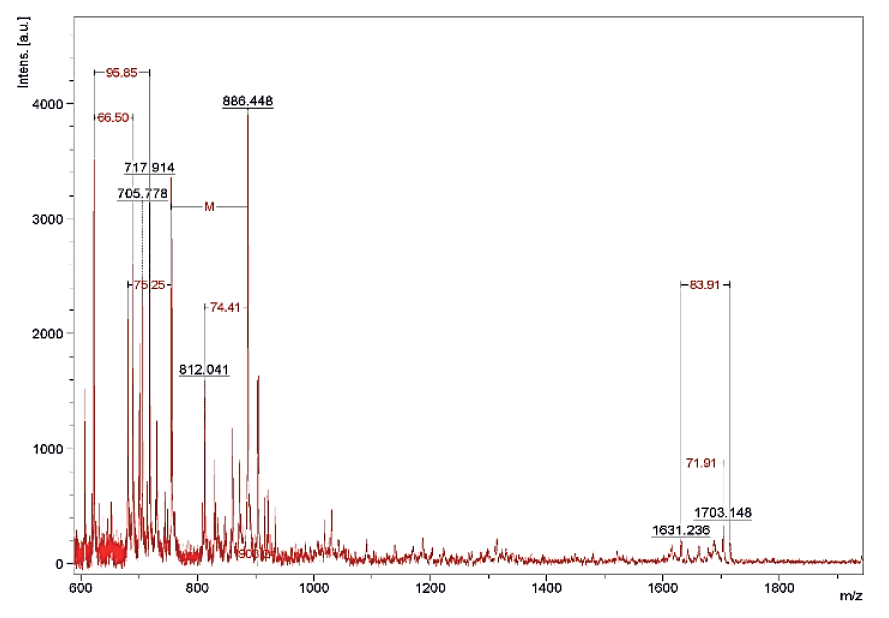

Having analyzed the mass spectra, we determined the molecular masses and amino acid sequences of seven peptides (Table 3).
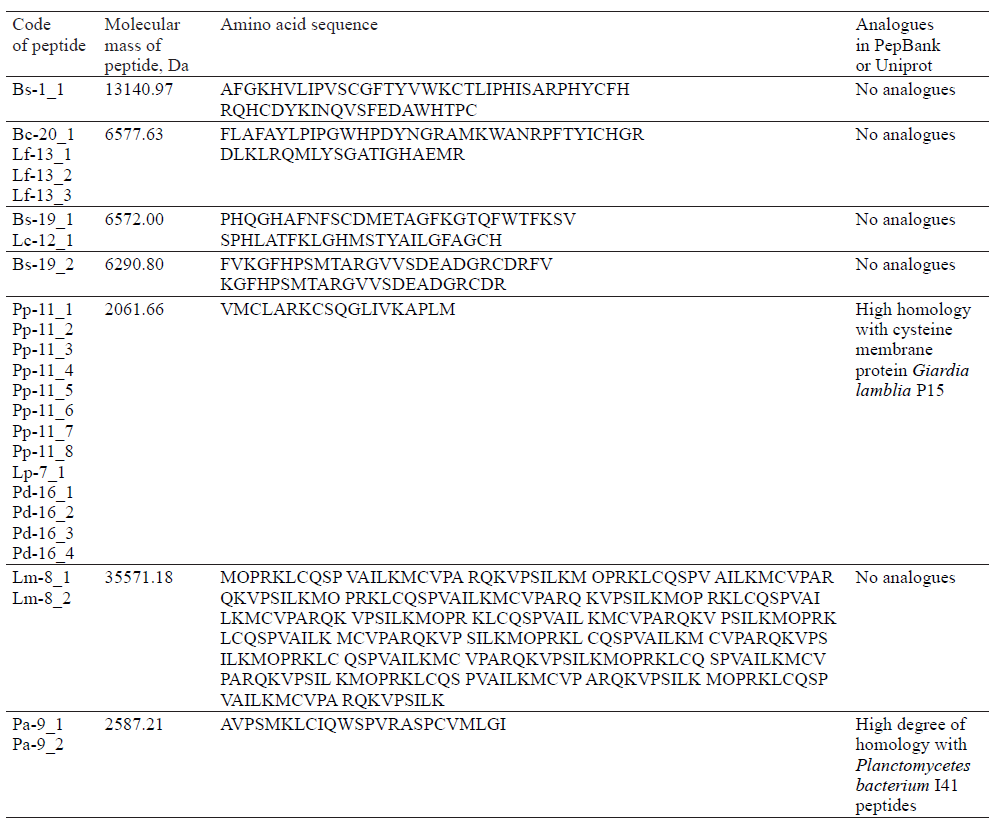
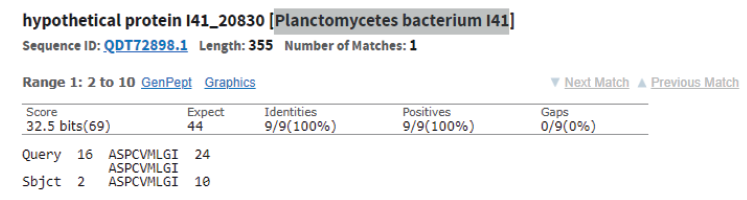
Table 3 also shows the presence of analogues for the studied peptides in the PepBank and Uniprot databases. We established a homology of fractions Pp-11_1, Pp11_2, Pp-11_3, Pp-11_4, Pp-11_5, Pp-11_6, Pp-11_7, Pp11_8, Lp-7_1, Pd-16_1, Pd-16_2, Pd-16_3, and Pd-16_4 with the cysteine membrane protein Giardia lamblia P15 (Fig. 8), as well as a homology of peptides Pa-9_1 and Pa-9_2 with the Planctomycetes bacterium I41 peptides (Fig. 9). The rest of the peptides had no analogues in the PepBank and Uniprot databases.

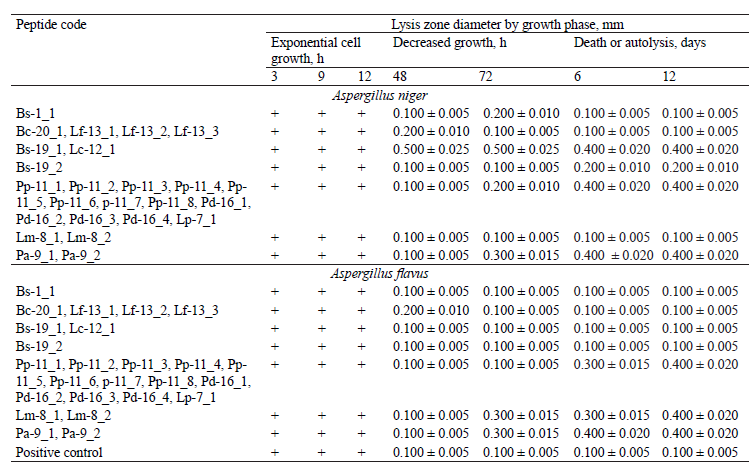
The antibacterial properties of the studied peptides against gram-positive (Bacillus pumilus) and gramnegative (Escherichia coli) bacteria, as well as their fungicidal properties against the microscopic fungi Aspergillus niger and Aspergillus flavus are presented in Tables 4–5 and Figs. 10–11.



According to Table 4 and Fig. 10, of the seven peptides under study, only one (Bs-19_2) exhibited no antagonistic activity against E. coli and B. pumilus strains. Peptide fraction Pp-11_1 (and peptides with identical amino acid sequences Pp-11_2, Pp-11_3, Pp11_4, Pp-11_5, Pp-11_6, Pp-11_7, Pp-11_8, Pd-16_1, Pd-16_2, Pd- 16_3, Pd-16_4, and Lp-7_1) showed high antagonism against B. pumilus and pronounced antibacterial activity against E. coli. Peptides Bs-1_1 and Bc-20_1 (identical Lf-13_1, Lf-13_2, and Lf-13_3), Lm-8_1 (identical Lm-8_2), and Pa-9_1 (identical Figure 5 Mass spectrum of Pp-11_1 fraction (Pp-11_2, Pp-11_3, Pp-11_4, Pp-11_5, Pp-11_6, Pp-11_7, Pp-11_8, Lp-7_1, Pd-16_1, Pd-16_2, Pd-16_3, Pd-16_4) Figure 6 Mass spectrum of Lm-8_1 fraction (Lm-8_2) Pa-9_2) had moderate and pronounced antagonistic activity against B. pumilus, but no activity against E. coli. Finally, peptide Bs-19_1 (identical Lc-12_1) showed bacteriostatic activity only against E. coli.
Unlike biocidal properties, which do not depend on the pathogen growth phase and naturally decrease over time, fungicidal properties need to be determined at each stage of the fungus life cycle since fungal pathogens have a complex growth cycle. We found that the peptide fractions under study did not stop fungal growth, but only inhibited it, which was indicated by a change in the mycelium color. The results were analyzed with time intervals (3, 9, 12, 24, 48, 72 h, etc.) and by the fungus growth phase (stationary, accelerated growth, logarithmic), i.e., during the periods of exponential cell growth, decreased growth, and death or autolysis. The samples with filters impregnated with a nutrient medium were used as a control.
Having analyzed the peptides’ fungicidal activity (Table 5, Fig. 11), we identified those peptides which could inhibit Aspergillus growth, rather than stop it completely. They were Bs-1_1, Bc-20_1 (identical Lf-13_1, Lf-13_2, and Lf-13_3) and Bs-19_2, with a lysis zone diameter of 0.1–0.2 mm. The maximum fungicidal activity against A. niger (0.3–0.5 mm lysis zone) was demonstrated by peptides Bs-19_1 (identical Lc-12_1), Pp-11_1 (identical Pp-11_2, Pp-11_3, Pp-11_4, Pp-11_5, Pp-11_6, p-11_7, Pp-11_8, Pd-16_1, Pd-16_2, Pd-16_3, Pd-16_4, and Lp-7_1), and Pa-9_1 (Pa-9_2). The highest activity against A. flavus (0.3–0.4 mm lysis zone) was revealed by peptides Pp-11_1 (identical Pp-11_2, Pp-11_3, Pp-11_4, Pp-11_5, Pp-11_6, p-11_7, Pp-11_8, Pd-16_1, Pd-16_2, Pd-16_3, Pd-16_4, and Lp-7_1), Lm8_1 (identical Lm-8_2), and Pa-9_1 (identical Pa- 9_2).
Based on the study of antimicrobial activity, we selected peptides with maximum antibacterial (against B. pumilus) and fungicidal (against A. niger and A. flavus) properties: Pp-11_1 (identical Pp-11_2, Pp-11_3, Pp-11_4, Pp-11_5, Pp-11_6, p-11_7, Pp-11_8, Pd-16_1, Pd-16_2, Pd-16_3, Pd-16_4, and Lp-7_1 ), Lm8_1 (identical Lm-8_2), and Pa-9_1 (identical Pa-9_2).
Thus, the fact that peptides produced by microorganisms inhabiting the natural ecosystems of Kuzbass exhibit antagonistic activity against opportunistic strains opens up prospects for their use in the production of pharmaceutical substances with antimicrobial action, alternative to traditional antibiotics.
ВЫВОДЫ
We identified amino acid sequences and molecular masses of peptide fractions produced by bacteria (Lactobacillus, Leuconostoc, Pediococcus, Bacillus, and Geobacillus) isolated from the natural objects of the Siberian region (soil, rhizosphere of plants, bottom sediments of reservoirs, and plant waste). In total, we isolated 25 protein fractions, some with identical mass spectra. Thus, we obtained seven peptides with different amino acid sequences, five of which have no analogues in the PepBank and Uniprot databases. One of the peptides (VMCLARKCSQGLIVKAPLM, 2061.66 Da) was homologous to the cysteine membrane protein Giardia lamblia P15, and another one (AVPSMKLCIQWSPVRASPCVMLGI, 2587.21 Da) was homologous to the Planctomycetes bacterium I41 peptides.
The peptides obtained from the culture fluid of bacteria isolated from natural sources of the Siberian Federal District were analyzed for antibacterial properties against Bacillus pumilus and Escherichia coli. We identified one peptide that exhibited no antagonistic activity against either gram-negative or gram-positive bacteria. One peptide fraction showed high antibacterial properties against both B. pumilus and E. coli. One peptide was active against E. coli, but not against B. pumilus (gram-positive bacteria). Finally, four out of seven peptides under study exhibited moderate and pronounced antagonism against B. pumilus, but no antibacterial activity against E. coli.
Our study of the peptides’ antifungal activity revealed three peptides that could inhibit the growth of the microscopic fungi Aspergillus niger and Aspergillus flavus, without stopping it completely (0.1–0.2 mm lysis zone). Four peptide fractions showed high fungicidal activity against Aspergillus (0.3–0.5 mm lysis zone).
According to our results, antimicrobial peptides produced by bacteria isolated from the natural objects of the Siberian region can be used as promising agents in the production of pharmaceutical substances and drugs (after safety trials) to treat infectious diseases, such as gastrointestinal, respiratory, blood and skin, as well as fungal infections.
Вклад авторов
The authors are equally responsible for the research results and the manuscript.КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that there is no conflict of interest.ФИНАНСИРОВАНИЕ
The research was conducted on the premises of the Research Equipment Sharing Center of Kemerovo State University (KemSU), agreement No. 075-15-2021-694 dated August 5, 2021, between the Ministry of Science and Higher Education of the Russian Federation (Minobrnauka) and Kemerovo State University (contract identifier RF----2296.61321X0032).СПИСОК ЛИТЕРАТУРЫ
- Mbarga MJA, Desobgo SCZ, Tatsadjieu LN, Kavhiza N, Kalisa L. Antagonistic effects of raffia sap with probiotics against pathogenic microorganisms. Foods and Raw Materials. 2021;9(1):24–31. https://doi.org/10.21603/2308-4057-2021-1-24-31.
- Kasjanenko SM, Kasjanenko OI, Nagornaya LV, Yevstafieva VA, Melnychuk VV, Lukyanova GA, et al. Yeast-rich mannan fractions in duck cultivation: prospects of using. Foods and Raw Materials. 2020;8(2):337–347. http://doi.org/10.21603/2308-4057-2020-2-337-347.
- Dyshlyuk L, Babich O, Ivanova S, Vasilchenco N, Atuchin V, Korolkov I, et al. Antimicrobial potential of ZnO, TiO2 and SiO2 nanoparticles in protecting building materials from biodegradation. International Biodeterioration and Biodegradation. 2020;146. https://doi.org/10.1016/j.ibiod.2019.104821.
- Babich O, Sukhikh S, Pungin A, Ivanova S, Asyakina L, Prosekov A. Modern trends in the in vitro production and use of callus, suspension cells and root cultures of medicinal plants. Molecules. 2020;25(24). https://doi.org/10.3390/molecules25245805.
- Musin KhG. Antimicrobial peptides – a potential replacement for traditional antibiotics. Russian Journal of Infection and Immunity. 2018;8(3):295–308. (In Russ.). https://doi.org/10.15789/2220-7619-2018-3-295-308.
- Salishcheva OV, Prosekov AYu. Antimicrobial activity of mono- and polynuclear platinum and palladium complexes. Foods and Raw Materials. 2020;8(2):298–311. http://doi.org/10.21603/2308-4057-2020-2-298-311.
- Jung C-J, Liao Y-D, Hsu C-C, Huang T-Y, Chuang Y-C, Chen J-W, et al. Identification of potential therapeutic antimicrobial peptides against Acinetobacter baumannii in a mouse model of pneumonia. Scientific Reports. 2021;11(1). https://doi.org/10.1038/s41598-021-86844-5.
- Bahar AA, Ren D. Antimicrobial peptides. Pharmaceuticals. 2013;6(12):1543–1575. https://doi.org/10.3390/ph6121543.
- Salishcheva OV, Prosekov AYu, Dolganyuk VF. Antimicrobial activity of mononuclear and bionuclear nitrite complexes of platinum (II) and platinum (IV). Food Processing: Techniques and Technology. 2020;50(2):329–342. https://doi.org/10.21603/2074-9414-2020-2-329-342
- Koo HB, Seo J. Antimicrobial peptides under clinical investigation. Peptide Science. 2019;111(5). https://doi.org/10.1002/pep2.24122.
- Mahlapuu M, Hkansson J, Ringstad L, Björn C. Antimicrobial peptides: An emerging category of therapeutic agents. Frontiers in Cellular and Infection Microbiology. 2016;6. https://doi.org/10.3389/fcimb.2016.00194.
- Zhang L, Gallo RL. Antimicrobial peptides. Current Biology. 2016;26(1):R14–R19. https://doi.org/10.1016/j.cub.2015.11.017.
- Giuliani A, Pirri G, Nicoletto S. Antimicrobial peptides: an overview of a promising class of therapeutics. Central European Journal of Biology. 2007;2(1):1–33. https://doi.org/10.2478/s11535-007-0010-5.
- Li J, Koh J-J, Liu S, Lakshminarayanan R, Verma CS, Beuerman RW. Membrane active antimicrobial peptides: Translating mechanistic insights to design. Frontiers in Neuroscience. 2017;11. https://doi.org/10.3389/fnins.2017.00073.
- Aoki W, Ueda M. Characterization of antimicrobial peptides toward the development of novel antibiotics. Pharmaceuticals. 2013;6(8):1055–1081. https://doi.org/10.3390/ph6081055.
- Kumar P, Kizhakkedathu JN, Straus SK. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules. 2018;8(1). https://doi.org/10.3390/biom8010004.
- Band VI, Weiss DS. Mechanisms of antimicrobial peptide resistance in gram-negative bacteria. Antibiotics. 2014;4(1):18–41. https://doi.org/10.3390/antibiotics4010018.
- Juturu V, Wu JC. Microbial production of bacteriocins: Latest research development and applications. Biotechnology Advances. 2018;36(8):2187–2200. https://doi.org/10.1016/j.biotechadv.2018.10.007.
- Zouhir A, Hammami R, Fliss I, Hamida JB. A new structure-based classification of gram-positive bacteriocins. Protein Journal. 2010;29(6):432–439. https://doi.org/10.1007/s10930-010-9270-4.
- B´edard F, Hammami R, Zirah S, Rebuffat S, Fliss I, Biron E. Synthesis, antimicrobial activity and conformational analysis of the class IIa bacteriocin pediocin PA-1 and analogs thereof. Scientific Reports. 2018;8(1). https://doi.org/10.1038/s41598-018-27225-3.
- Yang Y, Babich OO, Sukhikh SA, Zimina MI, Milentyeva IS. Identification of total aromas of plant protein sources. Foods and Raw Materials. 2020;8(2):377–384. DOI: http://doi.org/10.21603/2308-4057-2020-2-377-384.
- Johnson EM, Jung YG, Jin YY, Jayabalan R, Yang SH, Suh JW. Bacteriocins as food preservatives: Challenges and emerging horizons. Critical Reviews in Food Science and Nutrition. (2018);58(16):2743–2767. https://doi.org/10.1080/10408398.2017.1340870.
- Franz CMAP, van Belkum MJ, Holzapfel WH, Abriouel H, Galvez A. Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbiology Reviews. 2007;31(3):293–310. https://doi.org/10.1111/j.1574-6976.2007.00064.x.
- Bu Y, Liu Y, Li J, Liu T, Gong P, Zhang L, et al. Analyses of plantaricin Q7 synthesis by Lactobacillus plantarum Q7 based on comparative transcriptomics. Food Control. 2021;124. https://doi.org/10.1016/j.foodcont.2021.107909.
- Ranganath BS, Sharmila T, Balasubramanyam BV. Optimisation of media for the growth and production of bacteriocin from Bacillus coagulans. International Journal of Innovative Research in Technology, Science and Engineering. 2015;1(4):109–114.
- Sharma D, Singh Saharan B. Simultaneous production of biosurfactants and bacteriocins by probiotic Lactobacillus casei MRTL3. International Journal of Microbiology. 2014;2014. https://doi.org/10.1155/2014/698713.
- Milentyeva IS, Asyakina LK, Dyshlyuk LS, Velichkovich NS, Ostroumov LA. Research of biotechnological properties of microorganisms-antagonists. International Journal of Pharmaceutical Research. 2020;12(4):3351–3360. https://doi.org/10.31838/ijpr/2020.12.04.457.
- Milentyeva IS, Le VМ, Kozlova OV, Velichkovich NS, Fedorova AM, Loseva AI, et al. Secondary metabolites in in vitro cultures of Siberian medicinal plants: Content, antioxidant properties, and antimicrobial characteristics. Foods and Raw Materials. 2021;9(1):153–163. https://doi.org/10.21603/2308-4057-2021-1-153-163.
- Dyshlyuk LS, Babich OO, Ostroumov LА, Wang X, Noskova SYu, Sukhikh SA. Psychrophilic microorganisms in natural sources of the kemerovo region: isolation and antimicrobial properties. Food Processing: Techniques and Technology. 2020;50(4):763–773. (In Russ.). https://doi.org/10.21603/2074-9414-2020-4-763-773.
