

Аннотация
Fat of game animals is a prospective raw material for bioactive additives. Before such a product enters the market, food science has to make sure it is safe for consumption.This research featured subcutaneous adipose tissue of brown bears tested with standard methods for microbiological safety indicators. The microbial properties were studied on liquid and solid nutrient media. Staphylococcus was profiled using a VITEK 2 Compact biochemical automatic analyzer and Gram-positive cards (Bio-Mérieux, France). The analysis followed the Technical Regulations of the Customs Union TR TS 021/2011 On Food Safety (December 09, 2011).
The microbial count for mesophilic aerobic and facultative anaerobic microorganisms was 1.5×103 CFU/g. The fat samples revealed no molds, yeasts, or Escherichia coli bacteria. Liquid and solid nutrient media made it possible to describe the qualitative profile and cultural properties of the bear fat microflora against pork fat, which served as control. The automatic system identified Gram-positive, coagulase-negative, and oxidase-positive Staphylococcus lentus and Staphylococcus sciuri. In line with the modern classification, they belong to the new genus of Mammaliicoccus, namely Mammaliicoccus sciuri.
Subcutaneous adipose tissue of brown bears needs to undergo a microbiological safety test before consumption. Bear fat requires additional research in order to become a safe raw material for food products and bioactive additives.
Ключевые слова
Brown bear fat, subcutaneous adipose tissue, microbiological indicators, Mammaliicoccus lentus, Mammaliicoccus sciuriВВЕДЕНИЕ
The Strategy for Improving the Quality of Food Products in the Russian Federation through 2030 introduces several ways to develop an independent national food quality management system. This document stresses that the technical regulations for certain food products need a system of food quality indicators. New quality indicators will make it possible to test new foods with unconventional plant and animal raw materials, as well as to develop new biologically active additives and functional foods [1–5].
Adipose tissue of game animals is a potential source of bioactive additives [6]. In traditional medicine, bear (Ursus arctos L.) fat is an excellent anti-burn ointment. It also treats various diseases of the upper respiratory tract and heals skin damage of various severity. Bear fat is applied to relieve back and joints pain symptoms. The Mongols consume it raw; the Yakuts use twicemelted bear fat to treat tuberculosis [7]. Bear fat also is a popular folk remedy against gastrointestinal diseases and atherosclerosis [8, 9]. Customers buy it mostly online, and no technical regulations have been developed for it so far. As a result, the market is full of bear fat products of unknown safety status.
Raw fat is most often obtained from hibernating game animals, e.g., bears, badgers, marmots, etc. It is divided into subcutaneous and visceral. Most animal fat deposits are subcutaneous and are located right under the skin. Visceral adipose tissue is scarce and envelops the internal organs of well-fed adult specimen.
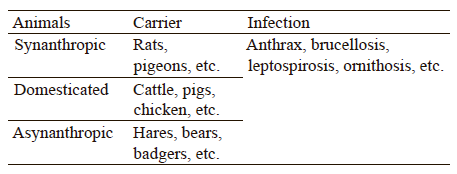
Meat and fat of farm and game animals can transmit zoonotic infections to people. Table 1 classifies them according to the carrier.
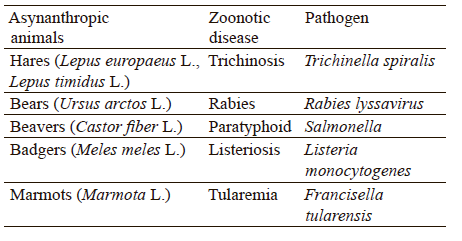
Game animals, being asynanthropic, are potential carriers of zoonotic diseases that can be transmitted to humans via infected meat and offal or via infected domestic and synanthropic animals. Table 2 describes the most common zoonoses of asynanthropic game animals.
Science knows more than a thousand pathogens of infectious diseases, 60% of which are of a zoonotic nature, i.e., people get them from animals. About 70% of such cases are connected with wild animals [10].
People usually get infected by inhaling contaminated aerosol secretions of rodents, e.g., feces, saliva, and urine. Meat and offal obtained from infected animals are also contagious. To be used as food, game meat and offal are to undergo a veterinary and sanitary examination, according to the Law on Veterinary Medicine of the Russian Federation (N 4979-1, May 14, 1993).
Meat and meat products contain saprophytic, sanitary-indicative, opportunistic, and pathogenic microorganisms. Microbial contamination occurs in endogenous and exogenous ways. Endogenous contamination takes place when the animal is still alive. Posthumous endogenous infection is usually associated with damaged intestines or ungutted carcasses. Exogenous contamination happens when microorganisms penetrate from the environment during butchering, storage, transportation, and processing. Retrocession from the gastrointestinal tract, i.e., endogenous infection of deep tissues in a living animal, occurs as a result of starvation, physical strain, diseases, and injuries. Microbial contamination in vivo is connected not only with the digestive system, but also with those systems that contact with the external environment, i.e., genitourinary, respiratory, and integumentary. Microbiotic studies of game animals are important precisely because the microflora of slaughter products depends on the composition of microorganisms the animal had when it was alive.
Bear microbiota is a popular subject of foreign studies. Bear intestinal microbiomes differ from those of other omnivores because bears have no caecum. The caecum restricts the rate at which nutrients pass through the intestinal tract. Apparently, it serves as a reservoir for microbial populations that replenish the microbiome diversity depending on the diet and health. Therefore, bears intestinal microbiomes are vulnerable to systemic changes in diet, health, or other factors. Gillman et al. believe that fecal samples provide insight into the intestinal microbiota of black bears, as well as other carnivores and omnivores with simple intestinal morphology [11].
Glad et al. studied the intestinal microbiome of polar bears and profiled microorganisms that belonged to the phylum of Firmicutes [12]. They identified 160 sequences as Clostridiales and found a new, unclassified sequence of Firmicutes. Most of the sequences (70%) belonged to Clostridium. The aerobic heterotrophic cell count on chocolate agar ranged from 5.0×104 to 1.0×106 CFU/mL for rectal swabs and from 4.0×103 to 1.0×105 CFU/g for feces samples.
Franz et al. studied intestinal microbiomes from two polar bear populations and identified eight most common classes of bacteria: Clostridia, Gammaproteobacteria, Actinobacteria, Coriobacteriia, Nogativicutes, Bacilli, Bacteroidia, Fusobacteria, Campylobacteria, and Saccharimonadia [13]. The microbiomes were different and reflected the habitat, diet, sex, and age of the animals. The authors decided that Bacilli were especially important for restoring intestinal health and maintaining intestinal homeostasis.
Schwab et al. studied fecal microbiota from ten grizzly bears [14]. The samples that belonged to wild grizzly bears contained more eubacteria than those obtained from captive bears. Enterococci and Enterobacteria were numerous in all samples. Pathogenic Clostridum perfringens group I had a positive correlation with protein intake and a negative correlation with dietary fiber content. Although considered healthy, the wild bears that lived on a normal protein-based diet were more likely to carry C. perfringens than those wild bears that relied mostly on plant-based food. Three samples even contained Clostridium sordellii, which can cause toxic shock syndrome in humans. Thus, the count of pathogenic Escherichia coli depended neither on the diet nor on the habitat.
Watson et al. and Trujillo et al. reported that subpopulations of brown bears living in different national parks shared five types of bacteria: Firmicutes, Proteobacteria, Epsilonbacteraeota, Bacteroidetes, and Actinobacteria [15, 16]. The authors identified 16 major genera. Therefore, bear intestinal microbiomes depend on the habitat, climate, age, sex, food diversity, and hunting strategy. Raw materials from game animals are to be checked for pathogenic and opportunistic pathogenic microflora before being processed into food products or biologically active additives.
Raw bear fat has a high lipid content, but it may also contain lipophilic microorganisms capable of synthesizing enzymes that hydrolyze lipids. Many bacteria, fungi, yeasts, and actinomycetes produce lipases that hydrolyze lipids at the water-fat phase boundary. Lipase-producing bacteria usually belong to Acinetobacter, Bacillus, Burkholderia, Pseudomonas, Staphylococcus, Microbacterium, Lactobacillus, Stenotrophomonas, Arthrobacter, Serratia, Aeromonas, Thermosyntropha, Achromobater, Chromobacterium, Burkholderia, Streptomyces, etc. [17–20].
Cooling, freezing, and thermal processing protect adipose tissue from bacterial contamination. Freezing and subsequent defrosting change the microbial quantitative and qualitative composition. Refrigerated storage gradually kills mesophilic microorganisms; however, some psychrophilic microorganisms remain viable for a long time.
Psychrotrophs proliferate on livestock products. Food-spoiling psychrotrophs are known to affect commercial foodstuffs. Zhang et al. identified microorganisms of 38 genera and 20 families, including Gramnegative bacteria [21]. Saprophytic Pseudomonas and especially Pseudomonas fragi had the highest count, followed by Psychrobacter, Brochothrix, Serratia, and Stenotrophomonas. Li et al. also reported other pathogenic and toxic microorganisms, such as Salmonella, Staphylococcus aureus, and C. perfringens [17]. Moschonas et al. detected psychrophilic and psychrotrophic anaerobic microflora in commercial Irish beef abattoir environments and vacuum-packed beef [22]. They tested 431 isolates and profiled 25 microbial species, with the most frequently recovered species being Clostridium gasigenes, followed by Clostridium estertheticum and Clostridium algidixylanolyticum. These species often cause spoilage in chilled lamb and vacuum-packed beef, which poses a significant commercial threat to the meat industry.
Pathogenic bacteria survive various methods of freezing and defrosting. Choi et al. studied the effect of freezing and defrosting on the microbiological quality and changes in the microstructure of chicken breasts inoculated with Listeria monocytogenes and Campylobacter jejuni [23]. They detected no differences in the count of L. monocytogenes u nder d ifferent f reezing conditions. However, air freezing (–20°C) reduced the total aerobic bacterial count and C. jejuni in particular, compared to other freezing methods. Defrosting by hot/cold air flow, water immersion, and high pressure at 4 and 25°C caused no significant differences in the count of L. monocytogenes.
Metzger et al. reported that foodborne pathogens survived freezing in cheese [24]. They produced three samples of semi-soft cheese with milk inoculated with two pathogen mixes of L. monocytogenes/S. aureus and E. coli/Salmonella typhimurium. Storage at –20°C for 2, 7, or 30 days resulted in little to no reduction in L. monocytogenes. However, 90 days at –20°C reduced the count of L. monocytogenes significantly, while the count of S. aureus remained constant over the 90-day storage in the freezer. E. coli and S. typhimurium rapidly decreased at –20°C. The defrosting conditions were 4°C for 14 h and 20°C for 4 h, but these factors had no effect on the viability of microorganisms.
Apparently, game meat and offal have to undergo a number of microbiological tests before consumption.
The research objective was to determine the microbiological safety profile and assess the qualitative composition of the microflora of bear adipose tissue.
The list of tasks included:
– reviewing scientific publications;
– developing a scheme for microbiological safety studies;
– analyzing microbiological safety criteria;
– isolating pure microbial cultures;
– profiling the isolated microflora; and
– comparing the obtained results with available publications.
The identified microbiological safety indicators can be used in technical regulations on quality and safety requirements for foods and dietary supplements based on bear fat.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
The research featured subcutaneous fat of Ursus arctos L. The samples belonged to a bear shot by a licensed hunter in the Kemerovo Region in 2021. The adipose tissue was separated from the carcass within 2 h after the slaughter. The butchering took place at –15 ± 5°C. For veterinary and sanitary examination, the samples were frozen at –18 ± 2°C and delivered to the laboratory within three days.
The Technical Regulations of the Customs Union TR TS 021/2011 On Food Safety contain no requirements for the microbiological safety of wild animal raw fat. Thus, we used the requirements for pork fat as control. The pork fat was purchased from the market and frozen under similar conditions.
After 12 h of freezing, the samples were defrosted at 20 ± 2°C in open air for 4 h, crushed in a cutter to a particle size of 3–5 mm, and packaged.
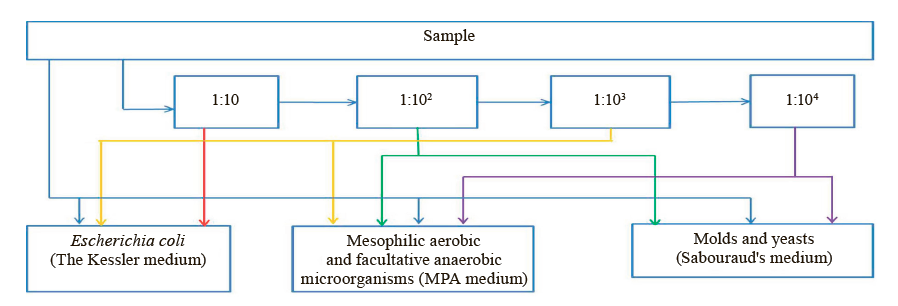
The sampling followed State Standard 31904-2012. We diluted 10 g of each sample in 90 cm3 of saline in the ratios of 1:10, 1:102, 1:103, and 1:104 by volume. Then, we dropped 1 cm3 of the substance with sterile pipettes in sterile Petri dishes and into test tubes with the Kessler medium. The procedure followed the microbiological control scheme illustrated in Fig. 1.
We profiled mesophilic aerobic and facultative anaerobic microorganisms in line with State Standard 10444.15-94. To study the QMAFAnM indicator, we used sterile meat peptone agar. After the nutrient medium solidified, Petri dishes were placed in a thermostat for cultivation at 37°C. After 48 h of cultivation, we counted the colonies and tested the indicators for the compliance with the requirements.
The procedure for Escherichia coli followed State Standard 31747-2012. We put the diluted product into a test tube with the sterile Kessler medium and a float and stored it in a thermostat at 37°C for 24 h. A bubble in the float indicated the presence of E. coli. The samples were tested for compliance with the Technical Regulations.
The mold and yeast tests corresponded with State Standard 10444.12-2013. We poured Sabouraud’s sterile nutrient medium into Petri dishes. After it solidified, we put the Petri dishes in a thermostat for cultivation at 25°C and counted the colonies after 72 h of cultivation. The Staphylococcus aureus test was in line with State Standard 31746-2012.
The cultural property test involved liquid and solid nutrient media. Staphylococcus bacteria were profiled using a VITEK 2 Compact biochemical analyzer (Bio-Mérieux, France) and a VITEK 2 Gram-positive identification card. This automated system provides 24-h microbial profiling and antimicrobial susceptibility testing. The software compares the test responses with the standard responses for each organism or group of organisms. A score and percentage probability indicate how the observed responses match the typical responses for each organism with a 99% probability.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
The Technical Regulations of the Customs Union provide no safety indicators for raw bear fat, so we used the requirements for pork fat as indicative indicators. Although they do not standardize the content of mold fungi and yeast in pork fat, we studied these indicators to assess the qualitative composition of the microflora, as well as the chance that these microorganisms might contaminate fat.
The pork fat and the bear fat complied with the TR CU 021/2011 in terms of microbiological safety criteria for pork fat. Table 3 shows the indicators of defrosted fat samples.

Pork fat and raw bear fat have a favorable chemical composition for microbial growth. The lipid part is 92% for pork fat and 72% for bear fat; they also contain 2–17% of proteins and 4–5% of moisture, respectively. Microorganisms can use these components as nutrients.
The total bacterial contamination was consistent with the data published by Maduka et al., who linked the higher bacterial count in pork fat with its physical properties, i.e., mucous nature and high fat content [25].
To determine QMAFAnM for the samples grown on the media, we established the taxonomic affiliation of microorganisms based on cultural, morphological, and tinctorial properties. The microflora of raw bear fat was represented by Bacilli. The microorganisms could be natural microflora or contamination during butchering, transportation, and storage. Other studies on the intestinal microbiota of brown, black, and polar bears also reported Bacilli [11, 13, 15]. Their representatives are known to produce lipase [19].
The research revealed some cocci that could be opportunistic pathogens for humans. Figures 2 and 3 show the colonies and the microscopy of bacterial preparations isolated from bear raw fat.

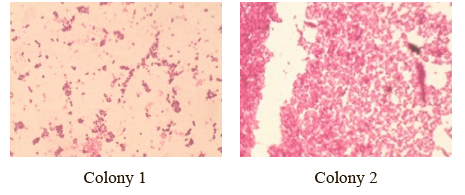
Colony 1 had a round shape with a diameter of 7 mm; it was white, flat, smooth, shiny, and opaque. Homogeneous in structure, it had a thick consistency, with serrated edges.
Colony 2 was round and slightly larger in diameter (11 mm). It was yellow, wavy, shiny, and opaque, with a thick and homogeneous consistency.
The microscopic image of the Gram-stained preparations made it possible to assess the shape and location of the microorganisms as cocci clustered like grape bunches. The cultural and morphological properties suggested that the microorganisms belonged to the genus of Staphylococcus.
Table 4 shows the identification of Staphylococcus isolated from the raw bear fat samples using the VITEK 2 Compact Bio-Mérieux automated system. The isolated microorganisms were identified as Grampositive, coagulase-negative, and oxidase-positive staphylococci consisting of clustered cocci.
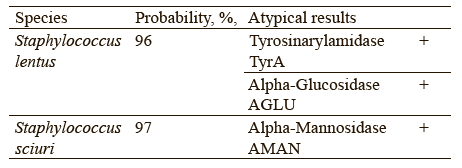
Chervyakova et al. proposed to include the following parameters into the list of authenticity markers: utilization of β-galactosidase and α-glucosidase, resistance to polymyxin B and novobiocin, ability to alkalinize lactate and N-acetyl-d-glucosamine [26]. These indicators could provide more accurate intraspecific profiling of Staphylococcus bacteria. Tables 5 and 6 give a detailed biochemical information on how the isolated microorganisms utilize particular components.
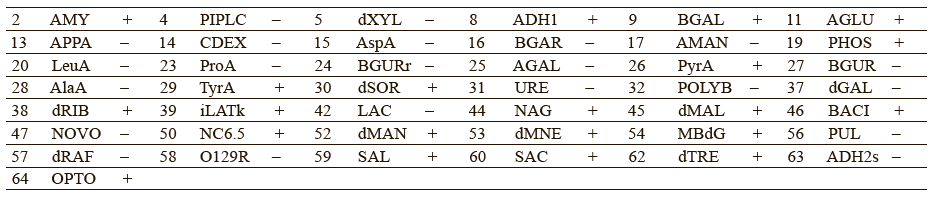
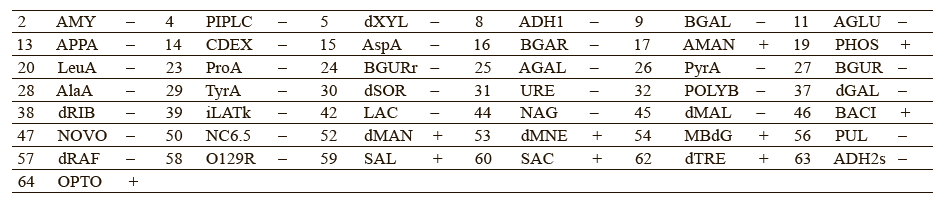
The experimental data suggested that Staphylococcus lentus and Staphylococcus sciuri belonged to the new genus of Mammaliicoccus, of which Mammaliicoccus sciuri is the type species. Staphylococcus fleurettii was also assigned to the new genus of Staphylococcus stepanovicii and Staphylococcus vitilinus [27].
M. sciuri and Mammaliicoccus lentus live on the skin and mucous membranes of many domestic, farm, and wild animals, as well as in foods of animal origin [28–33]. They occur in soil, sand, water, and marsh grass [34]. Adkins et al. found M. sciuri in milk and bedding on free-stall dairy farms [33]. M. sciuri were isolated from sick goats, cows with mastitis, dogs with dermatitis, cats with sepsis, and minks with urinary infections [35–37]. M. sciuri were also isolated from healthy and diseased humans [29].
ВЫВОДЫ
The microbiological safety of raw bear fat complied with the requirements for pork fat listed in Technical Regulation of the Customs Union TR CU 021/2011 On Food Safety. The VITEK 2 Compact Bio-Mérieux automated system detected Mammaliicoccus lentus and Mammaliicoccus sciuri in the samples grown on the QMAFAnM test medium. These microorganisms were reported as pathogens in animals.
The microbial profiling of bear adipose tissues indicated that foods and biologically active substances based on raw bear fat require safety tests, and their quality indicators have to be introduced into the existing regulatory documents.
Вклад авторов
E.A. Vechtomova and I.V. Dolgolyuk supervised the research, analyzed the data, interpreted the results, and wrote the article. M.M. Orlova and A.V. Zaushintsena reviewed scientific publications and performed the research.КОНФЛИКТ ИНТЕРЕСОВ
The authors declared no conflict of interests regarding the publication of this article.ФИНАНСИРОВАНИЕ
The research was supported by the Kemerovo State University (KemSU).СПИСОК ЛИТЕРАТУРЫ
- Sadovoy VV, Shchedrina TV, Trubina IA, Morgunova AV, Franko EP. Cooked sausage enriched with essential nutrients for the gastrointestinal diet. Foods and Raw Materials. 2021;9(2):345–353. https://doi.org/10.21603/2308-4057-2021-2-345-353
- Dubkova NZ, Kharkov VV, Vakhitov MR. Using Jerusalem artichoke powder in functional food production. Foods and Raw Materials. 2021;9(1):69–78. https://doi.org/10.21603/2308-4057-2021-1-69-78
- Tatarenko YaS, Koblov FS. Determination of the quality and safety of meat products. Achievements of Science and Technology in Agro-Industrial Complex. 2021;35(4):77–78. (In Russ.). https://www.elibrary.ru/VBLLBO
- Prosekov AYu. Scientific basis of food production. Kemerovo: Kemerovo Technological Institute of Food Industry; 2005. 21 p. (In Russ.). https://www.elibrary.ru/QNGNFL
- Vesnina A, Kozlova O, Prosekov A, Atuchin V. Genes and eating preferences, their roles in personalized nutrition. Genes. 2020;11(4). https://doi.org/10.3390/genes11040357
- Chernukha IM, Kupaeva NV, Smirnova JA, Akhremko AG, Pchelkina VA, Kotenkova EA. Methodology of adipose tissue type detection in mammals. Theory and Practice of Meat Processing. 2023;8(1):43–50. https://doi.org/10.21323/2414-438X-2023-8-1-43-50
- Nanzatov BZ, Sodnompilova MM. Folk medicine of Mongolian peoples: drugs of animal origin in beliefs and practices. Bulletin of the Irkutsk State University. Geoarchaeology, Ethnology, and Anthropology Series. 2016;17:126–145. (In Russ.). https://www.elibrary.ru/WXQQHZ
- Nasybullina NM. Animal fats as medicine. Meditsinskaya Sestra. 2012;(5):54–55. (In Russ.). https://www.elibrary.ru/PCGVJV
- Vechtomova EA, Kozlova OV, Orlova MM. Evaluation of methods for obtaining rendered animal fats. Food Processing: Techniques and Technology. 2022;52(4):797–806. (In Russ.). https://doi.org/10.21603/2074-9414-2022-4-2408
- Malov VA, Malova EYa. Zoonotic infections in modern society. Hidden threats. Medical Alphabet. 2016;3(19):42–43. (In Russ.). https://www.elibrary.ru/ZGIMWL
- Gillman SJ, McKenney EA, Lafferty DJR. Wild black bears harbor simple gut microbial communities with little difference between the jejunum and colon. Scientific Reports. 2020;10. https://doi.org/10.1038/s41598-020-77282-w
- Glad T, Bernhardsen P, Nielsen KM, Brusetti L, Andersen M, Aars J, et al. Bacterial diversity in faeces from polar bear (Ursus maritimus) in Arctic Svalbard. BMC Microbiology. 2010;10. https://doi.org/10.1186/1471-2180-10-10
- Franz M, Whyte L, Atwood TC, Laidre KL, Roy D, Watson SE, et al. Distinct gut microbiomes in two polar bear subpopulations inhabiting different sea ice ecoregions. Scientific Reports. 2022;12. https://doi.org/10.1038/s41598-021-04340-2
- Schwab C, Cristescu B, Northrup JM, Stenhouse GB, Gänzle M. Diet and environment shape fecal bacterial microbiota composition and enteric pathogen load of grizzly bears. PLoS One. 2011;6(12). https://doi.org/10.1371/journal.pone.0027905
- Watson SE, Hauffe HC, Bull MJ, Atwood TC, McKinney MA, Pindo M, et al. Global change-driven use of onshore habitat impacts polar bear faecal microbiota. The ISME Journal volume. 2019;13(12):2916–2926. https://doi.org/10.1038/s41396-019-0480-2
- Trujillo SM, McKenney EA, Hilderbrand GV, Mangipane LS, Rogers MC, Joly K, et al. Intrinsic and extrinsic factors influence on an omnivore's gut microbiome. PLoS One. 2022;17(4). https://doi.org/10.1371/journal.pone.0266698
- Li X, Boudjellab N, Zhao X. Combined PCR and slot blot assay for detection of Salmonella and Listeria monocytogenes. International Journal of Food Microbiology. 2000;56(2–3):167–177. https://doi.org/10.1016/S0168-1605(99)00209-3
- Adetunji AI, Olaniran AO. Production strategies and biotechnological relevance of microbial lipases: A review. Brazilian Journal of Microbiology. 2021;52(3):1257–1269. https://doi.org/10.1007/s42770-021-00503-5
- Sharma D, Sharma B, Shukla AK. Biotechnological approach of microbial lipase: A review. Biotechnology. 2011;10(1):23–40. https://doi.org/10.3923/biotech.2011.23.40
- Chandra P, Enespa, Singh R, Arora PK. Microbial lipases and their industrial applications: a comprehensive review. Microbial Cell Factories. 2020;19(1). https://doi.org/10.1186/s12934-020-01428-8
- Zhang Y, Wei J, Yuan Y, Yue T. Diversity and characterization of spoilage-associated psychrotrophs in food in cold chain. International Journal of Food Microbiology. 2019;290:86–95. https://doi.org/10.1016/j.ijfoodmicro.2018.09.026
- Moschonas G, Bolton DJ, McDowell DA, Sheridan JJ. Diversity of culturable psychrophilic and psychrotrophic anaerobic bacteria isolated from beef abattoirs and their environments. Applied and Environmental Microbiology. 2011;77(13):4280–4284. https://doi.org/10.1128/AEM.01778-10
- Choi EJ, Chung YB, Kin JS, Chun HH. Effects of freexing and thawing treatments on natural microflora, inoculated Listeria monocytogenes and Campylobacter jejuni on chicken breast. Journal of Food Hygiene and Safety. 2016;31(1):42–50. https://doi.org/10.13103/JFHS.2016.31.1.42
- Metzger N, Alvarez-Ordóñez A, Leong D, Hunt K, Jordan K. Survival of foodborne pathogens during frozen storage of cheese made from artificially inoculated milk. Dairy Science and Technology. 2015;95:759–767. https://doi.org/10.1007/s13594-015-0233-6
- Maduka CM, Oji A, Fineboy UQ, Okpokwasili GC. Microbial implications of beef fat and pork fat in the environment. Bionatura. 2020;5(4):1371–1374. https://doi.org/10.21931/RB/2020.05.04.15
- Chervyakova NS, Osin AV. Authentication of reference strains of pathogenic microorganisms applying automated microbiological analyzer “Vitek 2”. Problems of Particularly Dangerous Infections. 2017;(1):100–104. (In Russ.). https://doi.org/10.21055/0370-1069-2017-1-100-104
- Madhaiyan M, Wirth JS, Saravanan VS. Phylogenomic analyses of the Staphylococcaceae family suggest the reclassification of five species within the genus Staphylococcus as heterotypic synonyms, the promotion of five subspecies to novel species, the taxonomic reassignment of five Staphylococcus species to Mammaliicoccus gen. nov., and the formal assignment of Nosocomiicoccus to the family Staphylococcaceae. International Journal of Systematic and Evolutionary Microbiology. 2020;70(11):5926–5936. https://doi.org/10.1099/ijsem.0.004498
- Nemeghaire S, Argudín MA, Feßler AT, Hauschild T, Schwarz S, Butaye P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Veterinary Microbiology. 2014;171(3–4):342–356. https://doi.org/10.1016/j.vetmic.2014.02.005
- Couto I, Sanches IS, Sá-Leão R, de Lencastre H. Molecular characterization of Staphylococcus sciuri strains isolated from humans. Journal of Clinical Microbiology. 2000;38(3):1136–1143. https://doi.org/10.1128/JCM.38.3.1136-1143.2000
- Dakić I, Morrison D, Vuković D, Savić B, Shittu A, Ježek P, et al. Isolation and molecular characterization of Staphylococcus sciuri in the hospital environment. Journal of Clinical Microbiology. 2005;43(6):2782–2785. https://doi.org/10.1128/JCM.43.6.2782-2785.2005
- Stepanović S, Vuković D, Trajković V, Samardžić T, Ćupić M, Švabić-Vlahović M. Possible virulence factors of Staphylococcus sciuri. FEMS Microbiology Letters. 2001;199(1):47–53. https://doi.org/10.1111/j.1574-6968.2001.tb10649.x
- Klempt M, Franz CMAP, Hammer P. Characterization of coagulase-negative staphylococci and macrococci isolated from cheese in Germany. Journal of Dairy Science. 2022;105(10):7951–7958. https://doi.org/10.3168/jds.2022-21941
- Adkins PRF, Placheta LM, Borchers MR, Bewley JM, Middleton JR. Distribution of staphylococcal and mammaliicoccal species from compost-bedded pack or sand-bedded freestall dairy farms. Journal of Dairy Science. 2022;105(7):6261–6270. https://doi.org/10.3168/jds.2021-21500
- Barbaccia P, Gaglio R, Dazzi C, Miceli C, Bella P, Lo Papa G, et al. Plant growth-promoting activities of bacteria isolated from an anthropogenic soil located in Agrigento province. Microorganisms. 2022;10(11). https://doi.org/10.3390/microorganisms10112167
- Schauer B, Szostak MP, Ehricht R, Monecke S, Feßler AT, Schwarz S, et al. Diversity of methicillin-resistant coagulase-negative Staphylococcus spp. and methicillin-resistant Mammaliicoccus spp. isolated from ruminants and New World camelids. Veterinary Microbiology. 2021;254. https://doi.org/10.1016/j.vetmic.2021.109005
- Sacramento AG, Fuga B, Monte DFM, Cardoso B, Esposito F, Dolabella SS, et al. Genomic features of mecA-positive methicillin-resistant Mammaliicoccus sciuri causing fatal infections in pets admitted to a veterinary intensive care unit. Microbial Pathogenesis. 2022;171. https://doi.org/10.1016/j.micpath.2022.105733
- Schwarz S. Emerging chloramphenicol resistance in Staphylococcus lentus from mink following chloramphenicol treatment: characterisation of the resistance genes. Veterinary Microbiology. 1994;41(1–2):51–61. https://doi.org/10.1016/0378-1135(94)90135-x
