

Аннотация
Marker-assisted technologies in the field of plant biotechnology have attracted great interest of scientists seeking to determine the genetic variety and improve specific characteristics of species. Among several types, molecular markers hold great promise due to their high efficiency, adequate accuracy, and good reproducibility. This review aimed to present different molecular markers used in genetic biodiversity studies of common food plants, including potato, corn, and tomato.We presented some of the most frequent molecular markers in terms of their methodologies, advantages, challenges, and applications. We also reviewed the latest advances in the genetic diversity studies of common food plants that contribute to agricultural activities.
According to latest progress, Simple Sequence Repeats, Sequence Characterized Amplified Region, and Single Nucleotide Polymorphism are the most common molecular markers in plant diversity studies due to their co-dominancy, high level of polymorphism, great reproducibility, and adequate specificity. Considering common food plants like potato, corn, and tomato, Simple Sequence Repeats and Single Nucleotide Polymorphisms provide detailed information about polymorphisms, resistance to pathogens or diseases, genome maps, and population dynamics. However, more research should be conducted to apply the latest and more efficient technologies, such as Next Generation Sequencing, Diversity Array Technologies, and omics, to the genetic diversity studies of plant species.
Within the scope of recent progress, this review has a strong potential in providing relevant material for further research. It can serve as a guide to adopt the latest and most efficient sequencing platforms for examining various plant species, primarily potato, corn, and tomato.
Ключевые слова
Molecular marker, genome sequencing, Amplified Fragment Length Polymorphism (AFLP), Single Nucleotide Polymorphism (SNP), Random Amplified Polymorphic DNA (RAPD), polymerase chain reaction (PCR), potato, corn, tomatoВВЕДЕНИЕ
Today, one of the most striking challenges that the world faces is the growth of human population at an unstoppable rate. According to the latest forecasts, the world population is expected to reach 9.7 billion in 2050 and 11.2 billion in 2100, if no action is taken [1]. Moreover, there is an increasing concern about global warming that threatens the natural habitat of living organisms. These and similar environmental and sociological problems point out the potential risks in the availability of food, energy, water, and agricultural resources in the near future. Most of these problems involve the depletion of food plant resources that will present a serious threat to human and animal nutrition. Traditional agricultural activities that could protect these plants are limited since most cultivable lands are already under cultivation or urbanized. Also, environmental pollution and stress factors, such as drought, cold, salinity, and metal toxicity, make it difficult to carry out crop production. Therefore, researchers have focused on the search for more sustainable and economic solutions through technological and scientific innovations [2, 3].
The conservation of plant biodiversity and the sustainable use of existing resources are among the top priorities for researchers in various fields. With the recent developments in genetic engineering techniques, genetic variability and biodiversity of food plants have become a subject of molecular biology applications [4]. In particular, molecular markers are the key tools that are currently available to identify, conserve, and improve plant species. They represent DNA sequences that show polymorphism between individuals or populations. This technology offers an excellent opportunity for preserving the existing plant species, understanding metabolic pathways, increasing biodiversity, improving plant resistance to disease and insects, and developing new hybrid crops.
Notably, the application of marker technologies is not affected by climatic and environmental conditions, unlike agricultural practices. Some of the molecular markers include Amplified Fragment Length Polymorphism (AFLP), Random Amplified Polymorphic DNA (RAPD), Sequence-Related Amplified Polymorphism (SRAP), Inter Simple Sequence Repeats (ISSR), Simple Sequence Repeats (SSRs), Sequence Characterized Amplified Region (SCAR), Restriction Fragment Length Polymorphism (RFLP), Single Nucleotide Polymorphism (SNP), Diversity Array Technologies (DArT), and Next Generation Sequencing (NGS) [5].
The application of molecular marker technologies to preserve plant diversity is now being recognized as a way of solving the problems of traditional agriculture and food security. There have been a large number of completed and ongoing projects aimed at integrating molecular marker technologies into plant biotechnology studies. They have focused on a wide range of plant species including medicinal, wild, and edible plants that are highly important for biodiversity and human population. However, edible food plants are gaining much more attention as the world’s population is increasing, climatic conditions are getting worse, and humanity is facing serious food shortages [6].
Thus, a systematic review of molecular marker technologies applied to food plants can serve as a promising guide to develop novel strategies for preserving and improving these species. In this context, we aimed to present various molecular methods used in genetic biodiversity studies of some common food plants, including potato, tomato, and corn. These plants are essential and economically important vegetables among various types of crop plants in many countries. Also, they are highly preferable in the fight against food scarcity due to their nutritious and calorie-dense properties [7]. First, we summarized some of the most frequent molecular markers, their methodologies, advantages, challenges, and applications. Then, we reviewed the latest advances in genetic diversity studies of common plants that can aid agricultural activities. To our knowledge, this is one of the first reports that brings together the key aspects of the most recent progress in applying molecular marker technologies to potato, tomato, and corn plants.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
This review aimed to bring together the existing academic literature regarding the use of various molecular methods in genetic biodiversity studies of common food plants. Using relevant keywords (including “genetic diversity” and “molecular marker technology”), we searched for English-language articles mainly published in 2017–2022 and indexed by three databases, namely Web of Science, Science Direct, and Scopus. The list of publications was limited to high-quality peer-reviewed journals and the references in the retrieved papers were also screened for relevant studies.РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Molecular marker technologies. In the past, traditional breeding approaches to genetic diversity offered significant contributions to the identification and development of plant species. Some of the traditional methods included backcrossing, hybrid, and mutation breeding. However, these attempts were highly timeconsuming and susceptible to environmental conditions. Marker technology was developed to overcome these limitations and involved morphological, biochemical, and molecular markers. Molecular marker technology has gained much more attention due to the progress in molecular biology techniques and the challenges of the other markers [3, 8, 9]. Molecular markers are employed to detect different variations derived from insertions, deletions, or duplications located on chromosomes. They have several advantages over traditional methods as they show high efficiency and accuracy in all tissues with good reproducibility. Also, they are not affected by environmental conditions and/or the growth stage of plants [10, 11].
Molecular markers are mainly divided into three categories depending on their detection techniques: polymerase chain reaction (PCR)-based, hybridizationbased, and DNA sequence-based markers (Fig. 1). These classes include a variety of specific markers: Restriction Fragment Length Polymorphism (RAPD), Amplified Fragment Length Polymorphism (AFLP), Single Nucleotide Polymorphism (SNP), Sequence Characterized Amplified Region (SCAR), Inter Simple Sequence Repeats (ISSR), and Restriction Fragment Length Polymorphism (RFLP). They have their own pros and cons, which will be detailed in the next section [9].
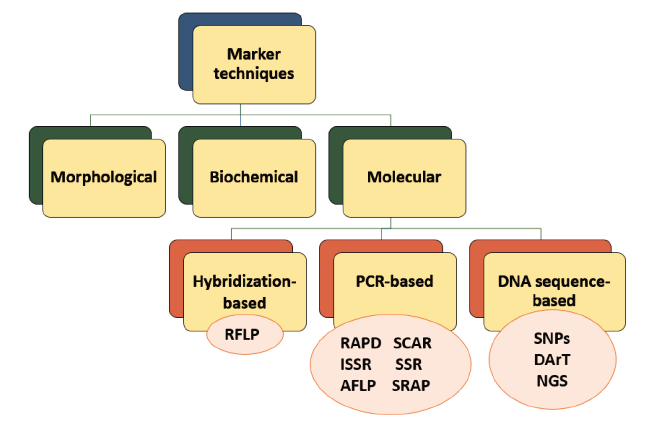
Hybridization-based marker techniques. The RFLP marker is the only method classified in hybridizationbased marker techniques. Changes in nucleotide sequences of the genomic DNA are derived from point mutations, insertions, deletions, translocations, duplications, etc., and they exhibit polymorphisms between individuals of species. These mutations can modify the restriction sites and lead to length variations in DNA fragments. When the genomic DNA is isolated and mixed with restriction enzymes, DNA is cleaved by these enzymes (restriction endonuclease, or restrictase) at cleavage sites. This is followed by the hybridization of the target DNA by a labeled probe after it is transferred to agarose gel electrophoresis and the Southern blotting.
RFLPs have been widely utilized as an efficient tool for the detection of polymorphisms, genome mapping, and population dynamic studies. For example, Kim et al. analyzed the quantitative trait locus of corn to determine its resistance to downy mildew, a major disease causing significant yield loss [12]. They used 691 SSR and 36 RFLP markers to identify polymorphisms between different lines, and reported that around one-third of the markers showed to be polymorphic. This technique has several advantages, including codominant inheritance, high reproducibility, locus specificity, and high genomic abundance. However, it requires a considerable amount of high-quality DNA, expensive toxic radioactive probes, and prior sequence information. Moreover, this method is time-consuming and costly [13, 14]. These disadvantages have guided researchers to develop other marker techniques.
PCR-based marker techniques. The PCR-based markers rely on the amplification of a particular DNA sequence in the presence of specific primers and enzymes by using the PCR technique [5, 15]. RAPD is one of the most common PCR-based marker systems that are generated with the random amplification of the genomic DNA in the presence of short arbitrary primers. After the PCR step, the separation and visualization of fragments by gel electrophoresis provide the detection of genomic polymorphisms by comparing the size of a band with the known molecular marker [16, 17]. Due to high speed, simplicity, and cost-effectiveness, the RAPD markers are highly preferred for a wide variety of plant species. For example, Sesli and Yegenoglu aimed to determine the effectiveness of RAPD and ISSR markers for the wild oil plant [18]. They reported that the RAPD marker provided reliable results about the genetic variability of this plant in a relatively shorter time than the ISSR marker.
In another study, RAPD, ISSR, and start codontargeted (SCoT) markers were compared to characterize the molecular profiles of five kalanchoe species. The authors found the RAPD marker to be considerably efficient in identifying the genetic polymorphism due to its high polymorphism level of 50–70% [19]. Notably, this marker requires a small amount (20–40 ng) of DNA and does not need any additional information prior to experiments [20, 21]. The major challenge of this technique is that the process and the obtained results are sensitive to slight changes in reaction conditions. Therefore, the RAPD protocol should be carried out under strictly controlled conditions. Otherwise, contradictions may be observed between different experiments resulting in lower reproducibility [21]. Sharma et al. compared the genetic diversity in carnation genotypes/mutants with RAPD, ISSR, and SSR markers [22]. They reported that the RAPD marker showed worse reproducibility than the other markers, since it was influenced by variable factors such as primer, template quantity, and amplification profile. One of the possible strategies to solve this problem was presented by Lin et al., who studied the development of a novel capillary electrophoresis-random amplified polymorphic DNA (CE-RAPD) method to improve the resolution of small molecular detection in RAPD profiling [23]. This method was defined as a promising tool for RAPD that provides standardization through repeated experiments.
The AFLP marker was developed to overcome the problems associated with the RAPD and RFLP methods. In detail, DNA is firstly digested by using two different restriction enzymes. Then, oligonucleotide adapters are ligated to the restriction fragments. Lastly, selective PCR is applied to amplify the attached fragments. Finally, the amplified fragments are checked by gel electrophoresis after they are stained with fluorescent or radioactive labelling [5].
AFLP is a dominant marker type that can use any DNA fragment regardless of its source, without requiring any prior data about the sequence. It can provide considerably reliable and reproducible results in a relatively shorter time than the RAPD and RFLP methods. Since restriction sites may be found across the whole genome, the AFLP markers can be used to simultaneously analyze a great number of polymorphisms with a single primer [24]. Therefore, this method plays a key role during the preparation of genetic and physical mapping. However, its application is limited because of some challenges, including its complexity. Moreover, it requires high molecular weight DNA and has a low ability to distinguish the homozygous from the heterozygous because of dominant characteristics. In addition to the method’s complexity, the observation of band profiles is difficult because of a short length of fingerprints in agarose gel [12, 25]. In order to resolve this, researchers have developed a novel Three Endonuclease-AFLP (TE-AFLP) method to analyze the genetic diversity of an Indian tea plant. As a result, clear banding profiles produced by the TE-AFLP technique were reported to be simple to score across gels [26].
SSRs, also called microsatellites, are a type of PCR-based markers which have short tandemly repetitive sequences found in the eukaryotic genome. Microsatellite DNAs have repeat motifs up to six nucleotides that show variations within and among individuals of a particular species. These variations and the length of sequences determine the level of polymorphism which can be detected with the PCR analysis.
The advantages of the SSR markers are high genomic abundance, co-dominant inheritance, ease of automation, high polymorphism, and moderate reproducibility. In comparison to the RAPD and AFLP markers, this method requires prior sequence information about the genome of a species for DNA sequence and it is a highly expensive procedure [16, 27]. As an alternative, ISSR markers are developed by modifying SSR, namely by amplifying genomic regions located between two identical repeat sections that are oppositely oriented. Although the ISSR markers are highly applicable in plant genetic studies, their limitations are low reproducibility and homology of co-migrating amplification products [5, 28]. Several studies have been performed to enhance the identification and authentication of plant species by using the ISSR marker technology. They revealed that ISSR coupled with the high resolution melting (HRM) analysis could provide beneficial results about the plant diversity and could be used to create new cultivars with desirable traits. The HRM analysis is a powerful technique that produces sequence-related melting profiles during dissociation of double stranded DNA and provides information about the differences in the genotype at the level of a single nucleotide [29, 30].
Co-dominant SCAR markers are an improved variant of RAPDs. They have higher specificity because the primers for SCARs are designed considering nucleotide sequences established in cloned RAPD fragments. Apart from high specificity, this marker system has such advantages as reproducibility, high efficiency with low quantities of a DNA template, ease of application, and low sensitivity to experimental conditions. Despite the numerous advantages, the use of SCARs is negatively affected by the need for additional information before the PCR and the detection limits of the primers. Regarding the detection limit, a wide variety of levels from 0.1 to 25 ng were reported as possible challenges. However, increasing the sample number and using the quantitative PCR may improve the detection limits [31, 32].
Another simple PCR-based marker is SRAP, a dominant and effective system for the production of genome-wide fragments. It is based on the amplification of open reading frames by utilizing two different primers, including CCGG sequence in the forward primer and AATT in the reverse primer. This marker is robust, efficient, and inexpensive. Thus, it is widely used in different applications, including map construction, genetic variety investigation, and DNA fingerprinting [17, 33].
DNA sequence-based marker techniques. The DNA sequencing technology has gained great attention with the developments in molecular biology. SNP is one of the sequence-based marker technologies that represents the most abundant and efficient DNA fingerprinting approach. This marker detects possible polymorphisms derived from changes in a single nucleotide position due to substitution, deletion, or insertion [13]. A variety of techniques have been reported to determine SNP genotypes, which are based on performing allelic distinction techniques and detection approaches. Some of the common detection methods are cleaved amplified polymorphic sequences (CAPS), Sanger sequencing, SNP-RFLP, and single strand conformational polymorphism (SSCP) [5]. The importance of SNP markers has risen with the latest developments in high-throughput genotyping methods like NGS, DArT marker, genotyping by sequencing, and allele-specific PCR.
DArT markers are microarray-based hybridization techniques that are largely used for mapping and genetic diversity studies. This system is considered a timesaving alternative to hybridization-based markers. DArT offers excellent potential to examine plant diversity, because it enables simultaneously genotyping of several thousands of polymorphic loci in a single assay. Also, it is highly reproducible and no prior information is needed to determine the sequence. The DArT marker analysis mainly involves the construction of a genetic library and its printing on microarray chips. After that, the labelled DNA is hybridized on chips and scanned for data analysis [5, 34]. The use of microarray platforms makes the DArT markers greatly preferred due to their ability to separate very high DNA fragment densities [35].
With the development of sequencing methods and high throughput technologies, simultaneous analysis of large amounts of DNA sequences has become an important approach. Currently, NGS techniques are becoming available for the sequencing of thousands to billions of nucleic acid sequences in one assay. They include a variety of techniques such as Illumina, Genapsys, Qiagen, Ion Torrent, and Roche 454 sequencing, which show different numbers of reads per run changing from 2 M to 1.1 billion. The massively parallel sequencing capacity of NGS provides more accurate analysis of the whole genome at a low cost [36, 37]. This technology has been widely applied to detect polymorphism, construct genetic maps, genotype mapping populations, and analyze whole genome sequences [17, 38]. However, the use of the NGS technology for plant species is still limited due to the lack of sufficient data for standard operating procedures. Also, the main challenges arise from the preparation of libraries since this technique generates massive amounts of data [39]. Therefore, there is a need for more research to accelerate plant diversity studies with the NGS technology.
Recent advances in genetic diversity of common food plants. Efforts have been made in the last years to protect some of the common food plants and improve their properties, such as resistance to pathogens, yield, and adaptation to environmental conditions. Applying genetic marker technologies to investigate plant diversity can help protect the existing plant sources and develop new strains with specific features [40, 41]. Some of the most recent molecular markers used to study variations in potato, corn, and tomato plants include RAPD, SSR, AFLP, SNP, SCAR, and DArT (Table 1). This review summarizes the latest developments in the genetic diversity studies of these three plants.
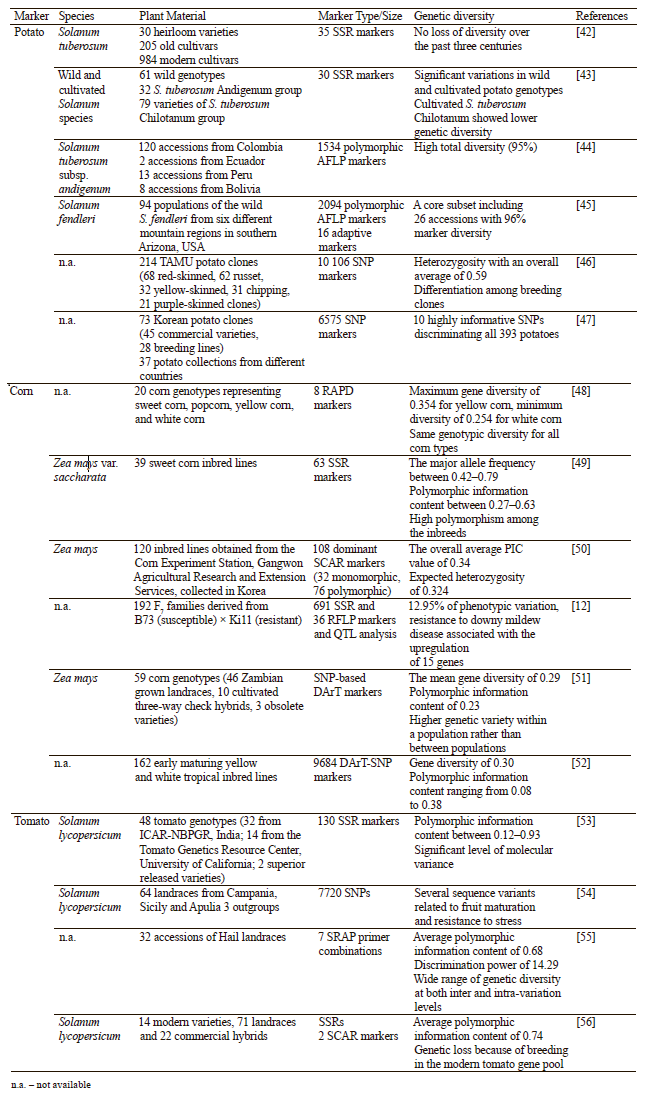
Genetic diversity of potato. Potato (Solanum tuberosum L.) is one of the most common food plants in the world, together with rice, corn, and wheat. It is a noncereal staple crop which serves as a major source for millions of people worldwide. China, India, and Russia are the leaders in potato production. In order to meet the global demand, it is crucial to improve its genetic potential [57]. For many years, scientists have studied different genetic varieties of potato using multiple marker-assisted technologies. Among them, SSRs have been mostly applied to characterize potato genetics due to their reproducibility, simplicity of use, and high polymorphism. A total of 1219 potato varieties from around the world have been investigated using SSRs to determine possible diversities over time and space. No major changes were reported in the genetic diversity of this crop over the past three centuries [42]. In another study, a diverse population of 189 genotypes of S. tuberosum was screened with SSR markers to determine the late blight resistance, which is one of the most destructive diseases in potato production. After analyzing wild and cultivated potato germplasms, the authors found significant resistance variations among the genotypes within the species [43]. These and similar studies (Table 1) have provided useful information for the determination of potato diversity and showed the effectiveness of SSR markers.
The AFLP marker technology is also used in potato genetic diversity studies. For example, del Rio and Bamberg aimed to analyze the minimum amount of germplasm units showing the highest plant diversity by building a core subset [45]. In their study, 144 accessions from the Potato Genebank located in the United States were discriminated from each other with genetic similarity values of 62–89%. After the selection process, a final core subset was built with a 96% marker diversity of 26 accessions.
The AFLPs are a key tool for potato studies because of high polymorphism and a small amount of DNA required. Yet, their use is limited by high cost. The DNA sequence-based markers, especially SNPs, can become an alternative to the PCR-based marker systems. Recently, Texas A&M University (TAMU) within its potato breeding program has investigated potato varieties and advanced clones at a molecular level by performing SNP markers. Their detailed analysis highlighted the genetic diversity of potato sources and paved the way for applying SNPs in other breeding programs [46]. Jo et al. carried out a similar study of potato clones from the Korean potato breeding program that provided useful information about the crop’s breeding history, regional adaptations, and market demands [47]. Despite the developments, the lack of sufficient data about potato proteomics and metabolomics makes it difficult to further investigate its genetic diversity and improve its quality.
Genetic diversity of corn. Corn (Zea mays L.) is another important crop worldwide that is consumed not only by humans but also by livestock. Its genetic diversity has been studied for many years and described in plentiful literature. Javed et al. applied the RAPD technology, one of the oldest marker systems, to assess different corn genotypes representing sweet corn, popcorn, yellow corn, and white corn [48]. According to their results, white corn had minimum gene diversity, while yellow corn showed the maximum level among 20 corn genotypes. Although some lines exhibited polymorphism with other groups, a total divergence was observed for all the studied types. As in potato diversity, the SSR markers have been commonly used to investigate the population structure and genetic diversities of corn [49, 58, 59].
In addition to the RAPD and SSR markers, SCARs, another PCR-based marker technology, may provide highly efficient data for the crop’s allele and genetic diversity. However, SCARs have not been applied in molecular studies of plants as commonly as the previous two markers. The potential of SCARs for corn diversity was investigated by Roy et al., who reported this marker’s effectiveness in analyzing the structure of corn inbred population of the Korean breeding lines [50].
Despite the great developments in sequencing technologies, recent efforts in corn studies have been shifted to the use of DNA sequence-based markers, and a number of successful results have been reported. One of the studies determined the genetic diversity of corn genotypes by using the SNP-based DArT marker [51]. In another study, 162 yellow and white corn inbred lines were analyzed for clarification of heterotic groups, intertrait relationships, and population structure with the use of the DArT-SNP markers [52]. These studies provided new insights into corn genetic diversity.
Genetic diversity of tomato. Tomato (Solanum lycopersicum L.), which originated in South America, is cultivated throughout the world as one of the most important vegetable crops. For example, the volume of tomato production in Russia is about 30 million tons. It has a high nutritional value due to a rich content of different metabolites such as lycopene, vitamins A and C, minerals, and dietary fiber. Therefore, researchers have used different technologies to study the genetic diversity of tomato and improve its properties.
The molecular markers that are generally used to explore tomato diversity are SSR, RAPD, SNPs, and ISSR [53, 60, 61]. For example, 48 genotypes of different exotic and indigenous sources were evaluated using 130 SSRs to determine the plant’s resistance to two common tomato diseases. The analysis resulted in developing a new species resistant to tomato spotted wilt virus and fusarium wilt diseases [53]. Gonias et al. employed characterization analyses by using SSRs coupled with SCAR, another reliable marker, to evaluate the polymorphism level of tomato germplasms [56]. This two-stage molecular approach produced useful data about the genetic variety and resistance of tomato to two serious fungal diseases, fusarium crown and root rot, as well as late blight. As a sequence based-marker, SNPs were used to analyze the genetic diversity of 64 tomato accessions from southern Italy. They showed a variety of mutations in genes, which were associated with stress tolerance and fruit quality [54]. Recently, the SRAP markers have been applied to analyze inter- and intra-genetic variability among Hail tomato landraces. The scientists determined the uniqueness of Hail tomato landraces and presented the potential use of the SRAP markers for other tomato-breeding programs which are of plant breeders’ interest to include in breeding programs for crop improvement. We assess the inter- and intra-genetic variability among 96 accessions representing three Hail tomato landrace using DNAbased marker sequence-related amplified polymorphism (SRAP) [55].
ВЫВОДЫ
Molecular marker technologies have found a wide range of applications in plant biotechnology because of their specific advantages in terms of efficiency, accuracy, reproducibility, simplicity, detection of polymorphism, and cost. This review summarized major molecular markers that have been used to study the genetic diversity of potato, corn, and tomato plants.
We identified different types of molecular marker technologies used for genetic diversity research of plant species. These markers are classified into three groups: hybridization-, PCR-, and sequence-based technologies. The PCR-based markers are more advantageous than the hybridization-based markers because of the latter’s toxicity and time-consuming characteristics. Considering the latest progress, SSRs and SCARs are highly preferred PCR-based markers due to their co-dominancy, high level of polymorphism, great reproducibility, and adequate specificity. However, rapid developments in molecular biology and biotechnology have led to a great shift towards sequence-based markers and novel marker technologies. One of such techniques are codominant SNPs, which show an extremely high level of polymorphism and good reproducibility. The recently developed NGS technology is a major breakthrough in the efforts to sequence plant genomes. This sequencing method has a strong potential to be used for plant diversity studies because it can provide reliable results by sequencing billions of nucleic acid sequences simultaneously. However, further research is needed to fully understand this technique and to validate its robustness for a variety of plant species.
Potato, corn, and tomato are essential and economically important vegetables, and their genetic information has been studied by several researchers. According to these studies, SSRs and SNPs are the most extensively used markers for these three food plants, since these markers possess high genomic abundance, co-dominant inheritance, ease of automation, high polymorphism, and adequate reproducibility. These markers provide detailed information about polymorphisms, resistance to pathogens or diseases, genome maps, and population dynamics. However, the most recent methodologies like DArT and NGS should be implemented into the genetic diversity studies of these plants to improve efficiency. This review can serve as a guide for adopting the latest and most efficient sequencing platforms to examine various plant species, primarily potato, corn, and tomato.
Вклад авторов
Bahar Aslanbay Guler conceptualized the research and wrote the original draft. Esra Imamoglu conceptualized and supervised the research, as well as reviewed and edited the manuscript.КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that there is no conflict of interest.СПИСОК ЛИТЕРАТУРЫ
- Sadigov R. Rapid growth of the world population and its socioeconomic results. Scientific World Journal. 2022;2022. https://doi.org/10.1155/2022/8110229
- Jones RAC, Naidu RA. Global dimensions of plant virus diseases: Current status and future perspectives. Annual Review of Virology. 2019;6:387–409. https://doi.org/10.1146/annurev-virology-092818-015606
- Ahmar S, Gill RA, Jung K-H, Faheem A, Qasim MU, Mubeen M, et al. Conventional and molecular techniques from simple breeding to speed breeding in crop plants: Recent advances and future outlook. International Journal of Molecular Sciences. 2020;21(7). https://doi.org/10.3390/ijms21072590
- Kersey PJ, Collemare J, Cockel C, Das D, Dulloo EM, Kelly LJ, et al. Selecting for useful properties of plants and fungi – Novel approaches, opportunities, and challenges. Plants, People, Planet. 2020;2(5):409–420. https://doi.org/10.1002/ppp3.10136
- Amiteye S. Basic concepts and methodologies of DNA marker systems in plant molecular breeding. Heliyon, 2021;7(10). https://doi.org/10.1016/j.heliyon.2021.e08093
- Cheng A, Mayes S, Dalle G, Demissew S, Massawe F. Diversifying crops for food and nutrition security – a case of teff. Biological Reviews. 2017;92(1):188–198. https://doi.org/10.1111/brv.12225
- Saeed F, Khan MA, Sharif M, Mittal M, Goyal LM, Roy S. Deep neural network features fusion and selection based on PLS regression with an application for crops diseases classification. Applied Soft Computing. 2021;103. https://doi.org/10.1016/j.asoc.2021.107164
- Lema M. Marker assisted selection in comparison to conventional plant breeding: Review article. Agricultural Research and Technology. 2018;14(2). https://doi.org/10.19080/ARTOAJ.2018.14.555914
- Hasan N, Choudhary S, Naaz N, Sharma N, Laskar RA. Recent advancements in molecular marker-assisted selection and applications in plant breeding programmes. Journal of Genetic Engineering and Biotechnology. 2021;19(1). https://doi.org/10.1186/s43141-021-00231-1
- Mondini L, Noorani A, Pagnotta MA. Assessing plant genetic diversity by molecular tools. Diversity. 2009;1(1):19–35. https://doi.org/10.3390/d1010019
- Singh HP, Raigar OP, Chahota RK. Estimation of genetic diversity and its exploitation in plant breeding. The Botanical Review. 2021;88:413–435. https://doi.org/10.1007/s12229-021-09274-y
- Kim HC, Kim K-H, Song K, Kim JY, Lee B-M. Identification and validation of candidate genes conferring resistance to downy mildew in corn (Zea mays L.). Genes. 2020;11(2). https://doi.org/10.3390/genes11020191
- Adhikari S, Saha S, Biswas A, Rana TS, Bandyopadhyay TK, Ghosh P. Application of molecular markers in plant genome analysis: a review. The Nucleus. 2017;60:283–297. https://doi.org/10.1007/s13237-017-0214-7
- Garrido-Cardenas JA, Mesa-Valle C, Manzano-Agugliaro F. Trends in plant research using molecular markers. Planta. 2018;247(3):543–557. https://doi.org/10.1007/s00425-017-2829-y
- Kumar A, Swapnil, Perween S, Singh RS, Singh DN. Prospect of molecular markers in precision plant breeding. In: Swapnil, editor. Recent advances in chemical sciences and biotechnology. New Delhi Publishers; 2020. pp. 131–142.
- Al-Hadeithi ZSM, Jasim SA. Study of plant genetic variation through molecular markers: An overview. Journal of Pharmaceutical Research International. 2021;33(45B):464–473. https://doi.org/10.9734/jpri/2021/v33i45B32828
- Nadeem MA, Nawaz MA, Shahid MQ, Doğan Y, Comertpay G, Yıldız M, et al. DNA molecular markers in plant breeding: current status and recent advancements in genomic selection and genome editing. Biotechnology and Biotechnological Equipment. 2017;32(2):261–285. https://doi.org/10.1080/13102818.2017.1400401
- Sesli M, Yegenoglu ED. Genetic relationships in wild olives (Olea europaea ssp. oleaster) by ISSR and RAPD markers. Biotechnology and Biotechnological Equipment. 2017;31(5):897–904. https://doi.org/10.1080/13102818.2017.1344107
- Al-Khayri JM, Mahdy EMB, Taha HSA, Eldomiaty AS, Abd-Elfattah MA, Latef AAHA, et al. Genetic and morphological diversity assessment of five kalanchoe genotypes by SCoT, ISSR and RAPD-PCR markers. Plants. 2022;11(3). https://doi.org/10.3390/plants11131722
- Tikendra L, Amom T, Nongdam P. Molecular genetic homogeneity assessment of micropropagated Dendrobium moschatum Sw. – A rare medicinal orchid, using RAPD and ISSR markers. Plant Gene. 2019;19. https://doi.org/10.1016/j.plgene.2019.100196
- Choudhury A, Deb S, Kharbyngar B, Rajpal VR, Rao SR. Dissecting the plant genome: through new generation molecular markers. Genetic Resources and Crop Evolution. 2022;69(1):2661–2698. https://doi.org/10.1007/s10722-022-01441-3
- Sharma P, Nath AK, Dhiman SR, Dogra S, Sharma V. Characterization of carnation (Dianthus caryophyllus L.) genotypes and gamma irradiated mutants using RAPD, ISSR and SSR markers. South African Journal of Botany. 2022;148:67–77. https://doi.org/10.1016/j.sajb.2022.04.012
- Lin W-J, Tung C-Y, Yen M-Y, Chan Y-J, Lin C-H, Hsueh P-R. A novel target pathogen identification and tracking system using capillary electrophoresis-random amplified polymorphic DNA. Scientific Reports. 2018;8(1). https://doi.org/10.1038/s41598-018-33702-6
- Ramesh P, Mallikarjuna G, Sameena S, Kumar A, Gurulakshmi K, Reddy BV, et al. Advancements in molecular marker technologies and their applications in diversity studies. Journal of Biosciences. 2020;45(1). https://doi.org/10.1007/s12038-020-00089-4
- Leipold M, Tausch S, Hirtreiter M, Poschlod P, Reisch C. Sampling for conservation genetics: How many loci and individuals are needed to determine the genetic diversity of plant populations using AFLP? Conservation Genetic Resources. 2020;12(4):99–108. https://doi.org/10.1007/s12686-018-1069-1
- Negi MS, Sharma SS, Singh A, Chauhan S, Adholeya A, Tripathi SB. Analysis of genetic diversity of Indian tea accessions using two modified amplified fragment length polymorphism methods. Proceedings of the National Academy of Sciences India, Section B: Biological Sciences. 2018;88:621–632. https://doi.org/10.1007/s40011-016-0798-8
- Li X, Qiao L, Chen B, Zheng Y, Zhi C, Zhang S, et al. SSR markers development and their application in genetic diversity evaluation of garlic (Allium sativum) germplasm. Plant Diversity. 2022;44(5):481–491. https://doi.org/10.1016/j.pld.2021.08.001
- Shahabzadeh Z, Mohammadi R, Darvishzadeh R, Jaffari M. Genetic structure and diversity analysis of tall fescue populations by EST-SSR and ISSR markers. Molecular Biology Reports. 2020;47(1):655–669. https://doi.org/10.1007/s11033-019-05173-z
- Buddhachat K, Changtor P, Ninket S. An accurate and rapid method for species identification in plants: Melting fingerprint-high resolution melting (MFin-HRM) analysis. Plant Gene. 2019;20. https://doi.org/10.1016/j.plgene.2019.100203
- Stavridou E, Lagiotis G, Kalaitzidou P, Grigoriadis I, Bosmali I, Tsaliki E, et al. Characterization of the genetic diversity present in a diverse sesame landrace collection based on phenotypic traits and EST-SSR markers coupled with an HRM analysis. Plants. 2021;10(4). https://doi.org/10.3390/plants10040656
- Ambreetha S, Balachandar D. SCAR marker: A potential tool for authentication of agriculturally important microorganisms. Journal of Basic Microbiology. 2022;63(1):4–16. https://doi.org/10.1002/jobm.202200419
- Dar AA, Mahajan R, Sharma S. Molecular markers for characterization and conservation of plant genetic resources. The Indian Journal of Agricultural Science. 2019;89(11):1764–1772. https://doi.org/10.56093/ijas.v89i11.95286
- Asande LK, Ombori O, Nyaboga EN, Oduor RO. Efficient shoot organogenesis using leaf disc and nodal explants of passion fruit (Passiflora edulis Sims) and genetic fidelity assessment using sequence-related amplified polymorphism (SRAP) markers. International Journal of Agronomy. 2020;2020 https://doi.org/10.1155/2020/3205710
- Loera-Sánchez M, Studer B, Kölliker R. DNA-based assessment of genetic diversity in grassland plant species: Challenges, approaches, and applications. Agronomy. 2019;9(12). https://doi.org/10.3390/agronomy9120881
- Emrey TM. Application of molecular markers SNP and DArT in plant breeding: A review paper. Journal of Agricultural and Crops. 2018;4(8):86–92.
- Nguyen KL, Grondin A, Courtois B, Gantet P. Next-generation sequencing accelerates crop gene discovery. Trends in Plant Science. 2019;24(3):263–274. https://doi.org/10.1016/j.tplants.2018.11.008
- Pervez MT, Hasnain MJU, Abbas SH, Moustafa MF, Aslam N, Shah SSM. A comprehensive review of performance of next-generation sequencing platforms. BioMed Research International. 2022;2022. https://doi.org/https://doi.org/10.1155/2022/3457806
- Manivannan A, Kim J-H, Yang E-Y, Ahn Y-K, Lee E-S, Choi S, et al. Next-generation sequencing approaches in genome-wide discovery of single nucleotide polymorphism markers associated with pungency and disease resistance in pepper. BioMed Research International. 2018;2018. https://doi.org/10.1155/2018/5646213
- Hess JF, Kohl TA, Kotrová M, Rönsch K, Paprotka T, Mohr V, et al. Library preparation for next generation sequencing: A review of automation strategies. Biotechnology Advances. 2020;41. https://doi.org/10.1016/j.biotechadv.2020.107537
- Soltabayeva A, Ongaltay A, Omondi JO, Srivastava S. Morphological, physiological and molecular markers for salt-stressed plants. Plants. 2021;10(2). https://doi.org/10.3390/plants10020243
- Munaweera TIK, Jayawardana NU, Rajaratnam R, Dissanayake N. Modern plant biotechnology as a strategy in addressing climate change and attaining food security. Agriculture and Food Security. 2022;11. https://doi.org/10.1186/s40066-022-00369-2
- Spanoghe M, Marique T, Nirsha A, Esnault F, Lanterbecq D. Genetic diversity trends in the cultivated potato: A spatiotemporal overview. Biology. 2022;11(4). https://doi.org/10.3390/biology11040604
- Duan Y, Duan S, Xu J, Zheng J, Hu J, Li X, et al. Late blight resistance evaluation and genome-wide assessment of genetic diversity in wild and cultivated potato species. Frontiers in Plant Science. 2021;12. https://doi.org/10.3389/fpls.2021.710468
- del Rio AH, Bamberg JB. Detection of adaptive genetic diversity in wild potato populations and its implications in conservation of potato germplasm. American Journal of Plant Science. 2020;11(10):1562–1578. https://doi.org/10.4236/ajps.2020.1110113
- del Rio A, Bamberg J. An AFLP marker core subset for the cultivated potato species Solanum phureja (Solanum tuberosum L. subsp. andigenum). American Journal of Potato Research. 2021;98(1):374–383. https://doi.org/10.1007/s12230-021-09849-w
- Pandey J, Scheuring DC, Koym JW, Coombs J, Novy RG, Thompson AL, et al. Genetic diversity and population structure of advanced clones selected over forty years by a potato breeding program in the USA. Scientific Reports. 2021;11. https://doi.org/10.1038/s41598-021-87284-x
- Jo KR, Cho S, Cho J-H, Park H-J, Choi J-G, Park Y-E, et al. Analysis of genetic diversity and population structure among cultivated potato clones from Korea and global breeding programs. Scientific Reports. 2022;12. https://doi.org/10.1038/s41598-022-12874-2
- Javed R, Iqbal M, Ullah S, Khan MR, Iqbal A, Ullah MS, et al. Phenotypic and molecular divergence in corn (Zea mays L.) ecotypes. Pakistan Journal of Agricultural Science. 2021;58:1777–1787.
- Mahato A, Shahi JP, Singh PK, Kumar M. Genetic diversity of sweet corn inbreds using agro-morphological traits and microsatellite markers. 3 Biotech. 2018;8(8). https://doi.org/10.1007/s13205-018-1353-5
- Roy NS, Park K-C, Lee S-I, Im M-J, Ramekar RV, Kim N-S. Development of CACTA transposon derived SCAR markers and their use in population structure analysis in Zea mays. Genetica. 2018;146(1):1–12. https://doi.org/10.1007/s10709-017-9985-7
- Kasoma C, Shimelis H, Laing MD, Shayanowako AIT, Mathew I. Revealing the genetic diversity of corn (Zea mays L.) populations by phenotypic traits and DArTseq markers for variable resistance to fall armyworm. Genetic Resources and Crop Evolution. 2021;68(4):243–259. https://doi.org/10.1007/s10722-020-00982-9
- Osuman AS, Badu-Apraku B, Ifie BE, Tongoona P, Obeng-Bio E, Garcia-Oliveira AL. Genetic diversity, population structure and inter-trait relationships of combined heat and drought tolerant early-maturing corn inbred lines from west and central africa. Agronomy. 2020;10(9). https://doi.org/10.3390/agronomy10091324
- Pidigam S, Thuraga V, Munnam SB, Amarapalli G, Kuraba G, Pandravada SR, et al. Genetic diversity, population structure and validation of SSR markers linked to Sw-5 and I-2 genes in tomato germplasm. Physiology and Molecular Biology of Plants. 2021;27(1):1695–1710. https://doi.org/10.1007/s12298-021-01037-8
- Tranchida-Lombardo V, Mercati F, Avino M, Punzo P, Fiore MC, Poma I, et al. Genetic diversity in a collection of Italian long storage tomato landraces as revealed by SNP markers array. Plant Biosystems – An International Journal Dealing with all Aspects of Plant Biology. 2019;153(2):288–297. https://doi.org/10.1080/11263504.2018.1478900
- Alzahib RH, Migdadi HM, Al Ghamdi AA, Alwahibi MS, Afzal M, Elharty EH, et al. Exploring genetic variability among and within hail tomato landraces based on sequence-related amplified polymorphism markers. Diversity. 2021;13(3). https://doi.org/10.3390/d13030135
- Gonias ED, Ganopoulos I, Mellidou I, Bibi AC, Kalivas A, Mylona PV, et al. Exploring genetic diversity of tomato (Solanum lycopersicum L.) germplasm of genebank collection employing SSR and SCAR markers. Genetic Resources and Crop Evolution. 2019;66(4):1295–1309. https://doi.org/10.1007/s10722-019-00786-6
- Ahmed S, Zhou X, Pang Y, Xu Y, Tong C, Bao J. Genetic diversity of potato genotypes estimated by starch physicochemical properties and microsatellite markers. Food Chemistry. 2018;257:368–375. https://doi.org/10.1016/j.foodchem.2018.03.029
- Belalia N, Lupini A, Djemel A, Morsli A, Mauceri A, Lotti C, et al. Analysis of genetic diversity and population structure in Saharan corn (Zea mays L.) populations using phenotypic traits and SSR markers. Genetic Resources and Crop Evolution. 2019;66:243–257. https://doi.org/10.1007/s10722-018-0709-3
- Iboyi JE, Abe A, Adetimirin VO. Microsatellite marker-based genetic diversity of tropical-adapted shrunken-2 corn inbred lines and its relationship with normal endosperm inbred lines of known heterotic classification. Plant Genetic Resources. 2020;18(6):454–461. https://doi.org/10.1017/S1479262120000489
- Herison C, Sutjahjo SH, Sulastrini I, Rustikawati R, Marwiyah S. Genetic diversity analysis in 27 tomato accessions using morphological and molecular markers. AGRIVITA. Journal of Agricultural Science. 2018;40(1):36–44. https://doi.org/10.17503/agrivita.v40i1.726
- Kiani G, Siahchehreh M. Genetic diversity in tomato varieties assessed by ISSR markers. International Journal of Vegetable Science. 2018;24(4):353–360. https://doi.org/10.1080/19315260.2017.1419397
