

Аннотация
Introduction. The fungus Aspergillus oryzae is widely used in the production of fermented soy-based products. However, there is little data on how its genetic characteristics affect the biochemical and fractional composition of protein substances during fermentation and the quality of fermented products. This study aimed to investigate the conversion of soy meal under the influence of two A. oryzae strains with different morphological and cultural properties during the production of a fermented soy sauce.Study objects and methods. The study used two A. oryzae strains, RCAM 01133 and RCAM 01134, which were isolated from the industrial F-931 strain (Russian Collection of Industrial Microorganisms), a producer of hydrolytic enzymes. Micromycetes were cultivated by a solid-phase method on soy meal, followed by dry fermentation. The results were analyzed with regard to accumulation of amine nitrogen, bound and free amino acids, proteins and carbohydrates.
Results and discussion. The cultivation of micromycetes resulted in a 35–38% increase in protein, a tenfold increase in free amino acids, and a 1.5–1.7 fold decrease in polysaccharides. The contents of essential amino acids in the fermented soy sauce were 1.7 and 1.2 times as high as in the initial medium (soy meal) and in the reference protein, respectively. Fermentation enhanced the biological value of proteins, increasing the amino acid scores of phenylalanine (7.3–7.7 times), phenylalanine (2 times), as well as valine, threonine, tryptophan, and lysine. The contents of protein and essential amino acids were slightly higher in the sauce with the RCAM 01133 strain.
Conclusion. Fermenting soy materials with the RCAM 01133 strain of A. oryzae is an alternative way to produce food ingredients with good sensory properties containing carbohydrates and biologically complete protein in easily digestible forms.
Ключевые слова
Micromycetes, soy meal, enzyme, protein, amino acids, amino acid scoreВВЕДЕНИЕ
Soy is the most common plant protein source in the world that has a wide range of health benefits. Clinical studies of soy-based products have revealed their antioxidant, antitumor, hypoglycemic, and hypotensive effects [1–5]. Having been cultivated since ancient times in Asian countries, since the 18th century in Europe, and since the 19th century in the USA, soy is now one of the most important agricultural crops worldwide [6, 7]. The USA is currently the leading producer of soybean, accounting for over 30% of world production. The popularity of soy products in North America has been growing over the past decades, especially after the Food and Drug Administration (FDA) linked soy protein with a reduced risk of coronary heart disease [8–10]. Soy protein contains all essential amino acids. Also, it has a higher biological value than plant proteins of cereals and a lower content of saturated fatty acids than animal products [11].
Asian countries mostly use fermented soy. Fermentation increases the solubility of soy polymers and improves absorption in the intestine, which provides soy-based products with high nutritional value and functional properties [12]. High soy consumption in Japan and China correlates with a low incidence of cardiovascular and oncological diseases. Soy-based products contain a large amount of soluble proteins, including bioactive peptides and amino acids, as well as isoflavones, polyamines, and other bioactive substances resulting from soy bioconversion [13–16].
Aspergillus oryzae is widely used in the production of fermented soy-based foods (soy sauce, soy paste, rice wine, etc.). The FDA declared the strains of this filamentous fungus safe for food production (GRAS), which was supported by the World Health Organization (WHO) [17–19]. The A. oryzae strains are industrial producers of enzyme preparations with proteolytic and amylolytic action [20]. The fungus synthesizes proteases as it adapts to a protein-rich environment during soy fermentation, with acidic proteases (carboxyl proteinase, carboxy-peptidase) playing the most important role in protein hydrolysis to form free amino acids [12, 20–23].
In addition, A. oryzae produces hydrolytic enzymes that break down polysaccharides into smaller monoand oligosaccharides, which can also affect the quality of fermented products [20, 24, 25]. Apparently, these enzymes play a regulatory role in the long-term conversion of plant materials, activating subsequent fermentation and supplying metabolites that act as precursors for the formation of the characteristic taste of soy sauce [26–29]. However, literature lacks information on the biosynthesis and catalysis of А. oryzae biomass or the influence of genetic and physiological characteristics of its strains on the biochemical and fractional composition of protein substances during microbial conversion of soy materials.
We conducted comparative studies of soy conversion under the influence of two A. oryzae strains with different morphological and cultural properties to produce a fermented sauce.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Our study objects were two strains of the filamentous fungus Aspergillus oryzae isolated as a result of selection from the F-931 industrial strain, a producer of proteolytic and amylolytic enzymes (Russian Collection of Industrial Microorganisms) [29]. The strains were deposited in the Russian Collection of Agricultural Microorganisms (RCAM) and differed in their morphological characteristics and cultural properties. RCAM 01133 and RCAM 01134 A. oryzae strains were used for the fermentation of soybean meal.
Micromycetes were cultivated by the solid-phase method in flasks on natural nutrient media containing soybean meal (55% moisture) for 4 days at 30–32°C. We used standard methods to analyze the activity of hydrolytic enzymes in the fungus biomass [30]. Microscopic studies were conducted with a Nikon Eclipse microscope (Japan).
Fungal cultures grown on soy were dry fermented for 48 h at 55–60°C in a TS-1/180 SPU dry-air thermostat. Surface cultures were extracted with a 20% NaCl solution in a ratio of 1:1.5 for 1 hour at 20–25°C. The incubation mixture was kept on a Biosan ES-20 (EU) incubator shaker at 200–210 rpm. Following the extraction, the soy sauce was separated in an OS-6M centrifuge at 5000 rpm for 10 min.
The biocatalytic conversion of fungal biomass polymers was evaluated according to the degree of accumulation of amine nitrogen, free amino acids, and general reducing substances [31]. The total protein content was determined by the Kjeldahl method on an automatic Vadopest system (Germany).
The amino acid content in the soy sauce samples was analyzed on a KNAUER EUROCHROM 2000 chromatograph. After that, the components were determined with a spectrophotometric Smartline UV Detector 2500 at λ = 570 nm (Germany). The aminograms were calculated by comparing the areas of the standard and the sample [32].
The amino acid score (AAS) was expressed as a percentage ratio between the amount of a certain essential amino acid in the test protein and that of the same amino acid in the FAO/WHO reference protein [33]:
![]()
where AAS is the amino acid score, %; M is the content of an essential amino acid in the test protein, g/100 g protein; MR is the content of an essential amino acid in the reference protein, g/100 g protein.
The experiments were carried out in at least three replicates. The data were statistically processed in Microsoft Excel using the Student’s coefficient (confidence interval 0.95).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
The solid-phase cultivation of Aspergillus oryzae RCAM 01133 and RCAM 01134 strains on nutrient media containing soybean meal revealed significant differences in their growth and development. At the first stage of growth (24 h), we observed swelling and germination of spores, as well as the formation of hyphae in both cultures.
At the second stage (42 h), RCAM 01133 showed more active mycelium formation accompanied by the release of metabolites and excess heat (the temperature in the medium layer increased to 34°C). RCAM 01134, however, featured a slightly slower rate of growth and mycelium formation.
By the third stage (72 h), both cultures had formed mycelium, which was airier and whiter in RCAM 01133. We also noticed the beginning of sporulation in RCAM 01134 (Fig. 1). The processes of biosynthesis and accumulation of enzymes took place at the same time, with the temperature of the medium dropping to 28–30°C.
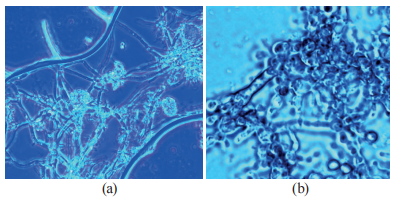
By the end of growth, on day 4, the fungal cultures looked like accreted colonies with 55–57% moisture. RCAM 01134 had more abundant yellow-green conidia, while RCAM 01133 had a white colony and a reduced ability to sporulate.
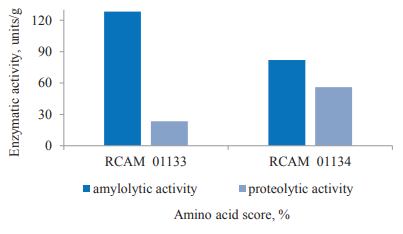
Then, we analyzed the biosynthetic features of the A. oryzae strains cultivated on soy. In particular, we measured the content of protein, polysaccharides, and amine nitrogen in surface cultures (Fig. 2), as well as determined the levels of proteolytic and amylolytic enzymes (Fig. 3).
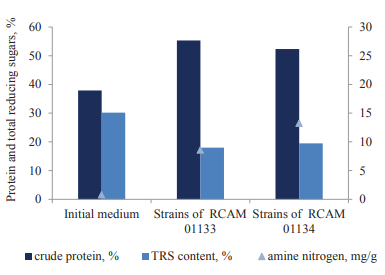
We found that the cultivation of the A. oryzae strains increased the content of crude protein by 35–38% and decreased the concentration of polysaccharides 1.5–1.7 times compared to the initial medium. The contents of protein and polysaccharides were 55.3% and 18.0% for RCAM 01133 and 52.3% and 19.5% for RCAM 01134, respectively. The content of amine nitrogen was 17 times as high in RCAM 01134 (13.24 mg/g) and 1.5 times as high in RCAM 01133 (8.59 mg/g), compared to the initial medium (0.8 mg/g).
The A. oryzae strains differed in the level of synthesized enzymes (Fig. 3). RCAM 01133 produced more α-amylase. The amylolytic activity of this strain reached 128.3 units/g, which was 1.6 times as high as that of the other strain. However, the proteolytic activity was 2.4 times higher in RCAM 01134 compared to RCAM 01133 (56.0 units/g and 23.5 units/g, respectively).
The grown fungal cultures were subjected to dry fermentation during which protein and carbohydrate polymers converted under the action of intracellular enzymes of micromycetes. After 48 h, the fermented biomass acquired a dark brown color, indicating the end of dry fermentation.
Then, we developed test samples of fermented sauces with the A. oryzae strains by extraction with saline. According to their biochemical and sensory analysis, the conversion of soy materials resulted in the accumulation of protein substances in the fermented sauces. The total amounts of amino acids were 23.0 g/100 g and 23.35 g/100 g in the sauces with RCAM 01133 and RCAM 01134, respectively. The percentages of essential amino acids amounted to 44.83% and 42.73%, respectively (Fig. 4.).
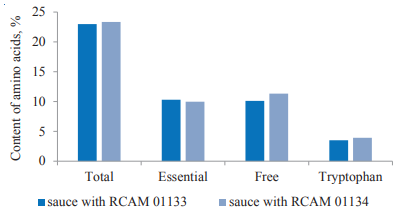
In addition to increasing total amino acids in the fermented sauces, caused by the synthesis of protein substances during the solid-phase cultivation of the fungus, the fermentation released amino acids from bound forms into free forms (Figs. 2, 4). Free amino acids accounted for 43.9% and 48.3% in the sauces with RCAM 01134 and RCAM 01134, respectively. About 50% of them were essential amino acids, which indicated higher medical and biological efficiency of the fermented products since free amino acids are easily absorbed into the blood, activating metabolic processes in the human body (Fig. 4).
The cultivation of A. oryzae micromycetes on a plant medium not only increased the content of protein substances but also improved their biological value in the fermented sauces. As we know, the content and composition of essential amino acids in the FAO/WHO reference protein can fully meet the physiological needs of the human body [34–36]. Our studies showed that the test samples of fermented soy sauce contained a full range of essential amino acids. Their amount was 1.7 times as high as in the original plant medium and 1.2 times as high as in the reference protein, mainly due to a high content of tryptophan (Table 1, Fig. 4).
We compared the composition of essential amino acids and their scores in the fermented products (Table 1, Fig. 5). The amino acid score, which is calculated by comparing the content of each amino acid in the proteins of the initial medium (soy meal) and the fermented sauces with its content in the reference protein, shows changes taking place during soy fermentation [37, 38].
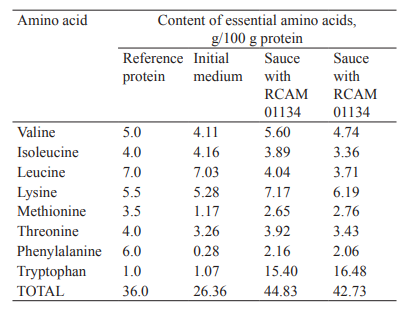
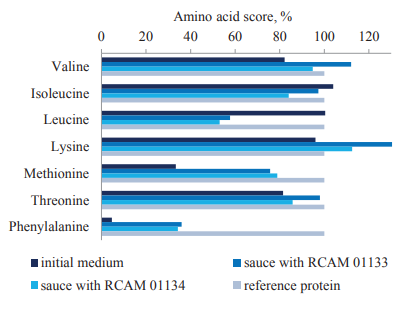
We found that phenylalanine, methionine, valine, and threonine were the limiting amino acids in the protein of the initial soy material, with their score under 82.2%. Although the score of the main limiting amino acid, phenylalanine, was only 4.7%, it increased 7.7–7.3 times in the fermented sauces, reaching 36.0–34.3%. The score of the second limiting amino acid, methionine, more than doubled as a result of fermentation. The scores also increased for valine, threonine, tryptophan, and lysine but decreased for leucine (Fig. 5).
The composition of essential amino acids in the fermented sauce proteins did not differ significantly, featuring three limiting amino acids – phenylalanine, leucine, and methionine. The score of the main limiting amino acids was slightly higher in the sauce with the RCAM 01133 strain of A. oryzae.
The increased score of individual amino acids (tryptophan, valine, lysine, methionine, and phenylalanine), which resulted from the fermentation of plant materials by micromycetes, indicated a higher biological value of the fermented products. The highest score was recorded for tryptophan, whose content was 15.40 g and 16.48 g per 100 g protein in the sauces with RCAM 01133 and RCAM 01134, respectively (Table 1).
It appears that the predominant amount of tryptophan gave the sauces their major functional properties. Tryptophan is known for its antidepressant, hypoglycemic, and cardioprotective effects. It helps to regulate appetite and synthesize the “joy hormone”, serotonin. Also, it stimulates the production of growth hormone and vitamin B3 (niacin). According to recent studies, tryptophan exerts a stress-protective effect on the cardiovascular system and acts not only as a precursor of the main neurotransmitter, serotonin, but also as a component of the body’s antioxidant system [39]. Tryptophan deficiency entails disturbances in the psycho-emotional state of a person such as sleep disturbances, digestive problems, and mental disorders. Together with magnesium deficiency, it can cause spasms of the coronary artery, cardiovascular pathology, and diabetes mellitus.
Increased amounts of valine, lysine, and methionine add more functional properties to fermented foods. They are among the most important essential amino acids, whose biological role in the human body is primarily associated with protein metabolism. They help with the growth and repair of tissues, accelerate cell regeneration, prevent the formation of lipoproteins, and participate in the regulation of the immune system [40, 41].
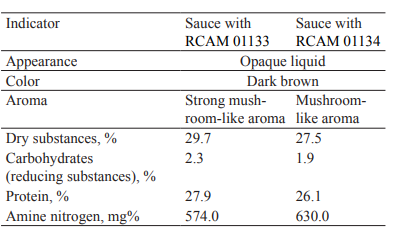
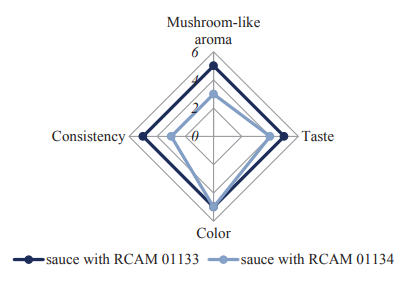
Sensory and physicochemical indicators of the fermented sauces are shown in Table 2 and Fig. 6. We found that the RCAM 01133-based sauce had a more pronounced mushroom-like aroma and a uniform consistency.
ВЫВОДЫ
Our study investigated the bioconversion of soy materials under the influence of two strains of the filamentous fungus Aspergillus oryzae that produce hydrolytic enzymes. The strains differed in their morphological characteristics and cultural properties.
We found that the cultivation of the RCAM 01133 and RCAM 01134 strains on a nutrient medium with soy meal increased the concentration of protein and amine nitrogen, as well as decreased the content of polysaccharides. The fermented soy sauce contained a full range of essential amino acids whose amount was larger than in the protein of the initial soy material and in the reference protein.
The comparative analysis of the fermented sauce proteins and the reference protein showed that the test samples had a higher biological value compared to the initial medium (soy meal). We found that fermentation increased the score of the main limiting amino acids, phenylalanine and methionine, as well as that of valine, threonine, tryptophan, and lysine. The score of leucine, however, decreased.
As a result, we selected the A. oryzae RCAM 01133 strain for use in biotechnology of functional additives based on microbial and biocatalytic conversion of plant materials. This strain had a high growth rate and a low spore-forming ability, which makes it technologically attractive for use in production.
To conclude, the fermented RCAM 01133-based soy sauce contained a full range of essential amino acids in free form and possessed good sensory properties. Thus, we can recommend it for ready meals as a seasoning and a salt substitute enriched with essential amino acids.
КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that there is no conflict of interest related to the publication of this article.ФИНАНСИРОВАНИЕ
The research was funded by the Ministry of Science and Higher Education of the Russian Federation (Minobrnauka) (No. 0529-2019-0066).СПИСОК ЛИТЕРАТУРЫ
- He FJ, Chen J-Q. Consumption of soybean, soy foods, soy isoflavones and breast cancer incidence: Differences between Chinese women and women in Western countries and possible mechanisms. Food Science and Human Wellness. 2013;2(3–4):146–161. https://doi.org/10.1016/j.fshw.2013.08.002.
- Allison DB, Gadbury G, Schwartz LG, Murugesan R, Kraker JL, Heshka S, et al. A novel soy-based meal replacement formula for weight loss among obese individuals: a randomized controlled clinical trial. European Journal of Clinical Nutrition. 2003;57(4):514–522. https://doi.org/10.1038/sj.ejcn.1601587.
- Zhang X-M, Zhang Y-B, Chi M-H. Soy protein supplementation reduces clinical indices in type 2 diabetes and metabolic syndrome. Yonsei Medical Journal. 2016;57(3):681–689. https://doi.org/10.3349/ymj.2016.57.3.681.
- Kim M-S, Kim B, Park H, Ji Y, Holzapfel W, Kim D-Y, et al. Long-term fermented soybean paste improves metabolic parameters associated with non-alcoholic fatty liver disease and insulin resistance in high-fat diet-induced obese mice. Biochemical and Biophysical Research Communications. 2018;495(2):1744–1751. https://doi.org/10.1016/j.bbrc.2017.12.003.
- Tutelʹyan VA, Pogozheva AV, Vysotskiy VG. Kliniko-gigienicheskie aspekty primeneniya soi [Clinical and hygienic aspects of the use of soy]. Moscow: Novoe tysyacheletie; 2005. 257 p. (In Russ.).
- Valliyodan B, Dan Q, Patil G, Zeng P, Huang J, Dai L, et al. Landscape of genomic diversity and trait discovery in soybean. Scientific Reports. 2016;6. https://doi.org/10.1038/srep23598.
- Li Y, Guan R, Liu Z, Ma Y, Wang L, Li L, et al. Genetic structure and diversity of cultivated soybean (Glycine max (L.) Merr.) landraces in China. Theoretical and Applied Genetics. 2008;117(6):857–871. https://doi.org/10.1007/s00122-008-0825-0.
- Huang H, Krishnan HB, Pham Q, Yu LL, Wang TTY. Soy and gut microbiota: Interaction and implication for human health. Journal of Agricultural and Food Chemistry. 2016;64(46):8695–8709. https://doi.org/10.1021/acs.jafc.6b03725.
- Velasquez MT, Bhathena SJ. Role of dietary soy protein in obesity. International Journal of Medical Sciences. 2007;4(2):72–82. https://doi.org/10.7150/ijms.4.72.
- Young VR. Soy protein in relation to human protein and amino acid nutrition. Journal of the American Dietetic Association. 1991;91(7):828–835.
- Sidorova YuS, Mazo VK, Kochetkova AA. Experimental evaluation of hypolipidemic properties of soy and rice proteins and their enzyme hydrolysates. A brief review. Problems of Nutrition. 2018;87(2):77–84. (In Russ.). https://doi.org/10.24411/0042-8833-2018-10021.
- Zorin SN, Vorob’eva IS, Vorob’eva VM, Netunaeva EA, Sidorova YuS, Kochetkova AA, et al. The processing of enzymatic hydrolysate of soy protein isolate. Food Industry. 2017;(8):13–15. (In Russ.).
- Okamoto A, Sugi E, Koizumi Y, Yanagida F, Udaka S. Polyamine content of ordinary foodstuffs and various fermented foods. Bioscience, Biotechnology and Biochemistry. 1997;61(9):1582–1584. https://doi.org/10.1271/bbb.61.1582.
- Galgano F, Caruso M, Condelli N, Favati F. Focused review: Agmatine in fermented foods. Frontiers in Microbiology. 2012;3. https://doi.org/10.3389/fmicb.2012.00199.
- Berezov TT, Makletsova MG, Fedorova TN. Polyamines: their role in normal condition and in disorders of the central neural systems. Annals of Clinical and Experimental Neurology. 2012;6(2):38–42. (In Russ.).
- Baraboy VA. Soybean isoflavones: biological activity and application. Biotechnologia Acta. 2009;2(3):044–054. (In Russ.).
- Tailor MJ, Richardson T. Applications of microbial enzymes in food systems and in biotechnology. Advances in Applied Microbiology. 1979;25:7–35. https://doi.org/10.1016/S0065-2164(08)70144-8.
- Abe K, Gomi K, Hasegawa F, Machida M. Impact of Aspergillus oryzae genomics on industrial production of metabolites. Mycopathologia. 2006;162(3):143–153. https://doi.org/10.1007/s11046-006-0049-2.
- FAO/WHO. Committee on Food Additives 31. Geneva: WHO; 1987.
- Rimareva LV, Serba ЕМ, Overchenko MB, Tadzhibova PYu, Serba EV, Krivova АYu, et al. Scientifically and experimental safety substitution of biotechnology products for the food processing industry. Vestnik of the Russian Agricultural Science. 2019;(1):40–43. (In Russ.). https://doi.org/10.30850/vrsn/2019/1/40-43.
- Liang Y, Pan L, Lin Y. Analysis of extracellular proteins of Aspergillus oryzae grown on soy sauce koji. Bioscience, Biotechnology and Biochemistry. 2009;73(1):192–195. https://doi.org/10.1271/bbb.80500.
- Xu D, Pan L, Zhao H, Zhao M, Sun J, Liu D. Breeding and identification of novel koji molds with high activity of acid protease by genome recombination between Aspergillus oryzae and Aspergillus niger. Journal of Industrial Microbiology and Biotechnology. 2011;38(9):1255–1265. https://doi.org/10.1007/s10295-010-0904-5.
- Zhao G, Hou L, Yao Y, Wang C, Cao X. Comparative proteome analysis of Aspergillus oryzae 3.042 and A. oryzae 100–8 strains: towards the production of different soy sauce flavors. Journal of Proteomics. 2012;75(13):3914–3924. https://doi.org/10.1016/j.jprot.2012.04.056.
- Machida M, Asai K, Sano M, Tanaka T, Kumagai T, Terai G, et al. Genome sequencing and analysis of Aspergillus oryzae. Nature. 2005;438(7071):1157–1161. https://doi.org/10.1038/nature04300.
- Wang B, Guo G, Wang C, Lin Y, Wang X, Zhao M, et al. Survey of the transcriptome of Aspergillus oryzae via massively parallel mRNA sequencing. Nucleic Acids Research. 2010;38(15):5075–5087. https://doi.org/10.1093/nar/gkq256.
- Feng Y, Cui C, Zhao H, Gao X, Zhao M, Sun W. Effect of koji fermentation on generation of volatile compounds in soy sauce production. International Journal of Food Science and Technology. 2013;48(3):609–619. https://doi.org/10.1111/ijfs.12006
- Lioe HN, Selamat J, Yasuda M. Soy sauce and its umami taste: a link from the past to current situation. Journal of Food Science. 2010;75(3):R71–R76. https://doi.org/10.1111/j.1750-3841.2010.01529.x.
- Serba EM, Mochalina PYu, Rimareva LV, Overchenko MB, Ignatova NI, Borshcheva YuA, et al. Fermented sauce based on microbial biomass. Food Industry. 2017;(9):28–30. (In Russ.).
- Serba EM, Overchenko MB, Rimareva LV, Pogorzhelskaya NS, Davydkina VE, Polyakov VA. Screening active population in fungus aspergillus oryzae ability to synthesize industrially significant metabolites. Mycology and Phytopathology. 2017;51(1):47–53. (In Russ.).
- Serba EM, Overchenko MB, Ignatova NI, Sokolova EN, Kurbatova EI. Development of national standards on methods for determining the activity of enzyme preparations. Food Industry. 2013;(7):40–44. (In Russ.).
- Polyakov VA, Abramova IM, Polygalina GV, Rimareva LV, Korchagina GT, Piskareva EN. Instruktsiya po tekhnokhimicheskomu i mikrobiologicheskomu kontrolyu spirtovogo proizvodstva [Instructions for techno-chemical and microbiological control of alcohol production]. Moscow: Deli print; 2007. 480 p. (In Russ.).
- Roslyakov VYa, Tarasenko IS, Balabanov NP, Vasilʹev PS. Opredelenie kolichestva aminokislot i peptidov v preparatakh parenteralʹnogo pitaniya na osnove gidroliza belka [Determination of the amount of amino acids and peptides in parenteral nutritional preparations based on protein hydrolysis]. Russian Journal of Hematology and Transfusiology. 1984;29(3):50–52. (In Russ.).
- Yushkov S. Razrabotka kompleksnogo sostava rastitelʹnykh belkov, imeyushchego polnotsennyy nabor aminokislot [Development of a complex composition of plant proteins with a complete set of amino acids]. Biznes pishchevykh ingredientov [Food ingredients business]. 2018;(1):22–27. (In Russ.).
- Protein and amino acid requirements in human nutrition. Report of a joint WHO/FAO/UNU expert consultation. WHO; 2007. 265 p.
- Dietary protein quality evaluation in human nutrition: Report of an FAO expert consultation. Rome: FAO; 2013. 79 p.
- Wolfe RR. The role of dietary protein in optimizing muscle mass, function and health outcomes in older individuals. British Journal of Nutrition. 2012;108:S88–S93. https://doi.org/10.1017/S0007114512002590.
- Zverev S, Nikitina M. Evaluation of protein quality in legumes. Kombikorma [Compound feed]. 2017;(4):37–41. (In Russ.).
- Nadtochiy LA, Orlova OYu. Innovatsii v biotekhnologii. CH. 2. Pishchevaya kombinatorika [Biotechnology innovation. Part 2. Food combinatorics]. St. Petersburg: ITMO University; 2015. 37 p. (In Russ.).
- Kratenko AS, Vovk KV, Sokruto OV, Nikolenko EYa, Alexandrova NK, Laricheva LV, et al. L-tryptophan: hypotensive, hypoglycaemic, cardioprotective effects and peculiarities of metabolism in modelled stress. Actual Problems of the Modern Medicine: Bulletin of Ukrainian Medical Stomatological Academy. 2017;17(1)(57):239–241. (In Russ.).
- Kalinin OV. Specific functions of essential amino acids. Molodezhʹ i nauka [Youth and science]. 2016;(1):2. (In Russ.).
- Bao X, Feng Z, Yao J, Li T, Yin Y. roles of dietary amino acids and their metabolites in pathogenesis of inflammatory bowel disease. Mediators of Inflammation. 2017;2017. https://doi.org/10.1155/2017/6869259.
