

Аннотация
Introduction. Probiotics are known for their beneficial properties. Numerous studies have been conducted to find advantages that probiotics can provide. This study aimed to evaluate the functional properties of raffia sap, a Cameroonian drink, fermented with probiotics by investigating its antagonistic activity against pathogenic bacteria.Study objects and methods. The study objective was raffia sap fermented by Lactobacillus fermentum and Bifidobacterium bifidum. Box-Behnken design with four factors (seeding rates of L. fermentum and B. bifidum, temperature, and incubation time) was used to generate mathematical models. The disc diffusion method was used to evaluate an antagonistic effect of the probiotics against four pathogenic bacteria (Escherichia coli, Listeria monocytogenes, Salmonella sp., and Bacillus cereus). An optimization of mathematical models of the inhibition diameters allowed to determine the optimal conditions of antagonistic effect.
Results and discussion. The experimental data showed that zones of inhibition were 0‒21 mm for Salmonella sp., 0‒23 mm for E. coli, 0‒20 mm for L. monocytogenes, and 0‒22 mm for B. cereus. ANOVA results and the mathematical models obtained showed that L. fermentum was effective against B. cereus and B. bifidum against Salmonella sp., E. coli, and B. cereus. The optimization of the models revealed maximum zones of inhibition at the seeding rates of L. fermentum and B. bifidum of 2 and 10%, respectively, incubation time of 48 h, and temperature of 37°C.
Conclusion. Raffia sap fermented by L. fermentum and B. bifidum demonstrated antagonistic effect against pathogenic bacteria such as E. coli, L. monocytogenes, Salmonella sp., and B. cereus.
Ключевые слова
Probiotics, antagonistic activity, pathogenic bacteria, response surface methodology, mathematical modelВВЕДЕНИЕ
Probiotics are defined as microorganisms that, when ingested in sufficient quantity, effect beneficially the host [1, 2]. The beneficial effects resulting from the consumption of foods enriched with probiotics have been known for millennia [3]. At the beginning of the 20th century, Mechnikov, a winner of the Nobel Prize, suggested replacing the dangerous germs by useful bacteria [4]. Additionally, Bifidobacterium spp. was recommended against infantile diarrhea [3, 5]. Despite the scientists’ research, the idea of eating certain bacteria to improve the health of the digestive system was ignored. Taking into account the different technical issues related to the production of foods with probiotics, attention must be focused on their beneficial effects on health [6].
The latest studies in this area have shown that probiotic bacteria are able to stimulate the immune system and inhibit the adhesion and multiplication of pathogenic bacteria [7, 8]. Since pathogenic microorganisms are becoming resistant to antibiotics, probiotics are a new alternative to be studied in the search for new molecules and/or antibacterial organisms [9].
Such an antimicrobial or antibacterial effect is generally called an antagonistic effect. Factors responsible for the antagonistic effect of one microorganism against another one are: production of organic acids or hydrogen peroxide that lower pH, competitive exclusion, immune system modulation, stimulation of defence systems, as well as production of antimicrobials such as bacteriocins and antioxidants [10]. Lactic, acetic, benzoic and other organic acids are the antimicrobial substances generally produced by beneficial microorganisms. The most produced bacteriocins are plantaricin, enterolysin, lacticin, lactocin, reuterin, pisciolin, enterocin, and pediocin [11].
Many probiotics have a broad spectrum of action and can be effective against diseases caused by food contaminated with certain pathogenic strains such as Listeria monocytogenes, Escherichia coli, Bacillus cereus and Salmonella. These four bacteria are the most common pathogens causing food-borne diseases [12]. Generally, there are difficulties in selecting an appropriate strain, substrate, as well as in determining optimal conditions for probiotic effectiveness.
In this context, researchers use local Cameroonian raw materials, including raffia sap. Raffia sap is a widespread drink in sub-Saharan Africa and particularly in Cameroon. Raffia sap undergoes wild fermentation and produces raffia wine that is difficult to keep. In 10 h after the harvest, alcohol produced during the primary fermentation transforms into acid, which seriously compromises the organoleptic characteristics appreciated by consumers.
In our previous research, we developed a probiotic beverage with raffia sap fermented by Lactobacillus fermentum and Bifidobacterium bifidum [13]. In the current research we studied an antagonistic potential of raffia sap inoculated by probiotics. The study was aimed to use Response Surface Methodology (RSM) to evaluate and optimize the effectiveness of L. fermentum and B. bifidum against E. coli, L. monocytogenes, Salmonella sp., and B. cereus.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Raffia sap harvesting. The fresh sap of less than eight hours was harvested in a 25 L container and transferred to the laboratory. The sap then was immediately dispensed into 1 L bottles and sterilized in a water bath at 65°C for 30 min. The bottles were cooled and stored at 4°C.
Bacteria and probiotics. Pathogenic bacteria (Escherichia coli, Listeria monocytogenes, Salmonella sp. and Bacillus cereus) were provided by the Food Microbiology Laboratory of Ngaoundere University. Probiotics (Lactobacillus fermentum and Bifidobacterium bifidum) were prepared using KwikStik™ lyophilized microorganism.
Revitalization and multiplication of probiotics. To revitalize and multiply probiotic cells contained in the freeze-dried products, 1 g of lyophilisate of each strain was rehydrated as recommended by the manufacturer. First, the powder was rehydrated in 10 mL of dilute saline solution (DS)consisting of 0.85% NaCl and 0.1% peptone in distilled water and stirred for 10 min until maximum recovery was reached. The solution was then transferred into 1 L of MRS broth previously prepared and sterilized. After incubation at 42°C for 48 h, MRS broth with probiotics was centrifuged at 6500 g and 4°C for 15 min.
The supernatant was removed, the pellet was washed in the saline solution without being resuspended and then recentrifuged as above. The supernatant was discarded and the pellet was finally resuspended in 10 mL of DS first and then transferred into 250 mL of DS. The concentration of probiotics in this solution was obtained by serial dilutions. The dilutions were spread on MRS petri dishes and incubated at 42°C for 24 h, then the colonies were counted [14].
Antagonistic effect of raffia sap fermented. To evaluate the antagonistic effect of the fermented raffia sap, we used the disc method described by Tadesse et al., with some modifications [5]. Mueller-Hinton agar was seeded with pathogenic bacteria (L. monocytogenes, B. cereus, E. coli and Salmonella sp.) and incubated at 37°C for 30 min. Sterile discs (5 mm) then were placed on the agar surface incubated at 37°C for 24 h. Each disk was impregnated with 100 μL of raffia sap fermented by probiotics according to the experimental design (Table 1). The inhibition of pathogenic bacteria resulted in the formation of clear zones around the discs. The zone of these inhibition zones was measured, which was used as the main response of the trial.
Experimental design for sap fermentation process and data analysis. Fermentation was done following a four factor Box-Behnken design. The factors were seeding rates of L. fermentum (X1) and B. bifidum (X2), temperature (X3), and incubation time (X4). The levels of each factor were chosen after prior testing (Table 1).
The Box-Benhken experimental matrix in coded variables (–1; 0; +1) was generated with the Minitab 18 software. This coded variable matrix consisted of 28 trials, four of which enabled a better evaluation of the experimental error; each trial was repeated three times. The experimental matrix applicable to the laboratory was obtained by transforming the matrix into the coded variables with the EXCEL software using the following formula:
![]()
where Xj is a value of the coded variable j; Uj is a value of the real variable j; Uj0 is a value of real variable j at the center, and ΔUj is called a “step” of variation.
Modelling and optimization. The zones of inhibition zones obtained after the application of the various tests of the experimental matrix were analysed on Minitab 18. The obtained models were in the form of:
![]()
where y is the model of the inhibition zones of the strain concerned, β(i,j) are model coefficients and x(i,j) are the factors. The data was analysed at the level of 10%, including the maximum of significant factors on each model. Response Surface Methodology was used for the three-dimensional graphical representation of the models of each inhibition zone after setting temperature and incubation time at constant values. Sigmaplot 12 software was used to plot the curves. Optimization was done on Minitab with the specifications for maximizing the inhibition zones of each pathogen.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
The results of the measurements of the inhibition zones of pathogenic bacteria (Salmonella sp., Escherichia coli, Listeria monocytogenes, and Bacillus cereus) obtained after the implementation of the four factor Box-Behnken experimental matrix showed that the zones of inhibition ranged from 0 to 21 mm for Salmonella sp., 0 to 23 mm for E. coli, 0 to 20 mm for L. monocytogenes, and 0 to 22 mm for B. cereus.
Raffia sap without probiotics did not demonstrated an inhibitory activity against pathogenic bacteria (inhibition zone = 0). However, the study conducted by Ojo and Agboola displayed different results [15]. The authors evaluated the antagonistic activity of bacteria isolated from Palm wine (Raphia vinifera L.) towards Salmonella typhi. The study revealed that raffia sap, due to its own microbial flora, was antagonistic against several pathogenic bacteria, including Salmonella sp. This also could be explained by pasteurization of fresh sap to avoid any interaction between the natural microflora of the sap and added probiotics, as well as wild fermentation.
Thus, seeding rates of Lactobacillus fermentum and Bifidobacterium bifidum played an important role in the antagonistic effect of the drink against the pathogenic bacteria tested, but statistical analysis was performed for a better demonstration of these effects (Table 2).
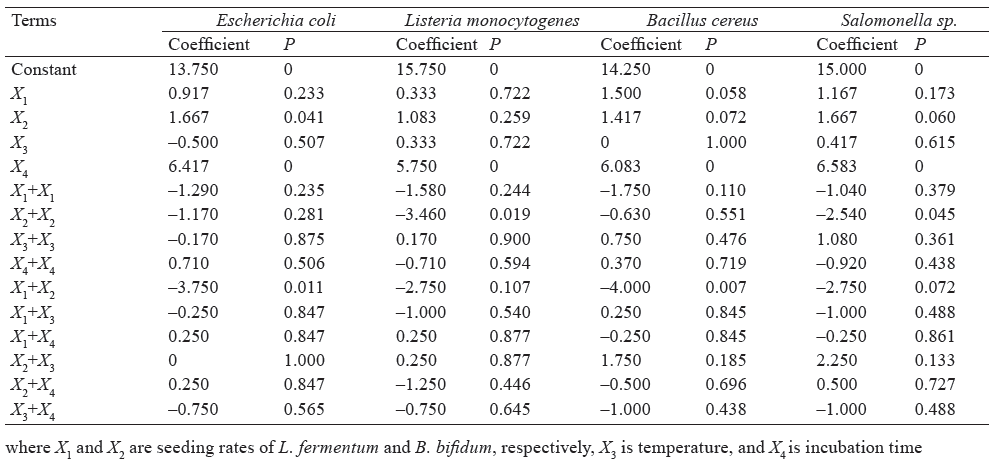
Effect of factors on microbial inhibition. According the data in Table 2, B. bifidum did not show a strong antagonistic effect on E. coli, L. monocytogenes, and Salmonella sp., but it was effective against B. cereus (P ≤ 0.1). L. fermentum had a significant antagonistic effect on Salmonella sp., E. coli, and B. cereus with probabilities of 0.060, 0.040 and 0.072, respectively. Moreover, the incubation time significantly increased all the zones of inhibition (P = 0.000).
Effect of incubation time on inhibition of pathogenic bacteria. The curves of inhibition zone of E. coli, L. monocytogenes, B. cereus, and Salmonella sp. as a function of time were obtained after fixing seeding rates of B. bifidum and L. fermentum at 0 in coded variables (5% in real variables) and the temperature at 0 in coded variable (39.5°C in real variable). Under these conditions, these curves (Fig. 1) showed that the inhibition zones of E. coli ranged from 8 mm (2 h of incubation) to 20 mm (48 h).

The inhibition zones of B. cereus, L. monocytogenes, and Salomonella sp. varied from 8.5, 9.0 and 7.3 mm in 2 h of incubation, respectively. In 48 h, the zones reached 20 mm in all the samples. The inhibition zones measured for each pathogenic strain as a function of the incubation time demonstrated that time is an essential factor to assess the antagonistic effect of probiotic drink based on raffia sap fermented with L. fermentum and B. bifidum. In fact, B. bifidum and L. fermentum need time to synthesize acids and other antimicrobial compounds contributing to antagonist effect against pathogenic bacteria [16, 17].
Individual effect of B. bifidum on E. coli, B. cereus and Salmonella sp. To obtain the inhibition curves of E. coli, B. cereus, and Salmonella sp. (Fig. 2) as a function of the seeding rate of B. bifidum, the seeding rate, incubation temperature, and incubation time of L. fermentum were set at 0 in coded variable ‒ 5%, 39.5°C, and 25 h in real variables, respectively.
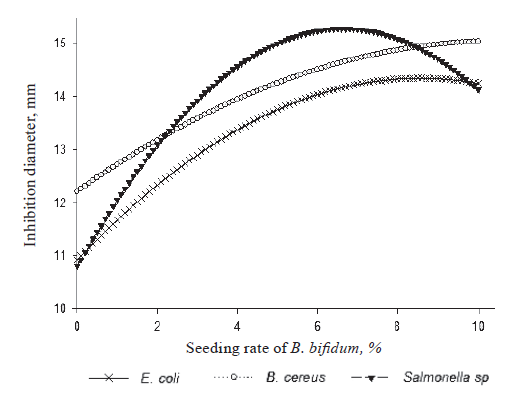
The inhibition curve of Salmonella sp. as a function of the seeding rate of B. bifidum showed that the maximum zone of inhibition of Salmonella sp. (15 mm) was obtained when the seeding rate of B. bifidum was 6%. The curve of the inhibition zone of B. cereus demonstrated that the inhibition zone depended directly on the seeding rate of B. bifidum. The inhibition zones of B. cereus ranged from 12.1 to 14.2 mm for the seeding rates of 0 and 10%. As for E. coli, its curve of the inhibition increased and then decreased, with a peak of 13.3 mm when the seeding rate of B. bifidum was 6.6 %.
According to Luquet and Corrieu, bifidobacteria promote better absorption of milk lactose in adults with intestinal lactase deficiency [18]. In our study, these probiotics (in particular B. bifidum) in raffia sap also played an important antagonistic role against E. coli, B. cereus, and Salmonella sp. In addition, some invitro studies showed that bifidobacteria and their metabolites stimulated IgA production, phagocytic activity, and growth [19]. These metabolites produced in raffia sap as well as the B. bifidum strain itself can therefore be a natural way to stimulate the immune system, to inhibit pathogenic strains such as E. coli, B. cereus and Salmonella sp., and to balance intestinal flora.
Individual effect of L. fermentum on B. cereus. Figure 3 shows the curve of the inhibition zone of B. cereus as a function of the seeding rate of L. fermentum. This curve increased then decreased, with the inhibition zone peak of 14.3 mm at the seeding rate of 6.5%. This curve was obtained by setting the seeding rate of B. bifidum, incubation temperature, and incubation time at 0 in coded variables ‒ 5%, 39.5°C, and 25 h in real variables, respectively.
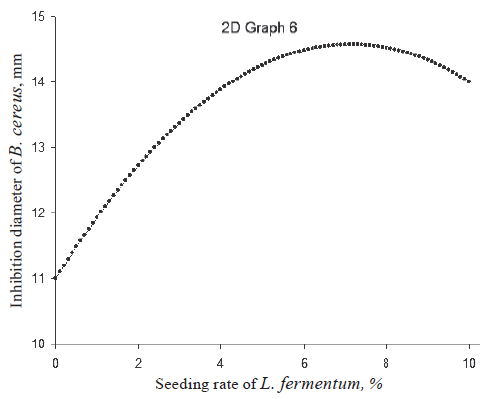
Thus, if it were necessary to optimize the antagonistic properties of our probiotic drink by referring only to an ability to inhibit the B. cereus strain, the seeding rates of L. fermentum and B. bifidum would be 5% and 5%, respectively, with an incubation temperature of 39.5°C and an incubation time of 25 h. Under these conditions, this probiotic drink could eventually be used as a means of combating infectious diseases which can be caused by B. cereus. B. cereus is a group of bacteria that can be pathogenic for humans. The infections they can cause are generally infrequent and not serious. However, ingestion of these bacteria, and their toxins in particular, can lead to infections characterized by vomiting or diarrhea [20].
In spite of the fact that our results were obtained in vitro, it is clear that L. fermentum introduced into raffia sap had a significant antagonistic effect on B. cereus. However, further research should be carried out in vivo to take into account factors that could affect the drink properties such as its passage through the intestinal tract, the survival of strains and the bioavailability of antibacterial compounds, as well as their direct or indirect effect on the body.
Effects produced by combination of L. fermentum and B. bifidum in raffia sap on the pathogens tested. The response surface methodology was applied to represent the mathematical models obtained by holding temperature and incubation time at 0 in coded variables ‒39.5°C and 25 h in real values, respectively.
Figure 4 presents the response surface of the mathematical model of inhibition zone of L. fermentum and B. bifidum against Salmonella sp. An increase in the seeding rate of B. bifidum and a simultaneous increase in the seeding rate of L. fermentum and B. bifidum considerably increased the antagonistic effect, with the inhibition zone of 16 mm.
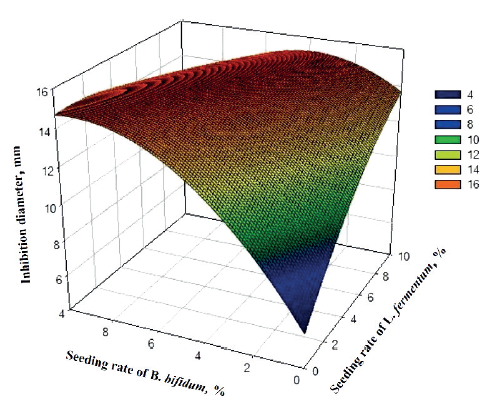
However, only B. bifidum had a significant antagonistic effect on Salmonella sp. (P = 0.060, Table 2) at a 10% probability level. Indeed, lactic acid produced by B. bifidum lowers the pH by creating an unfavorable conditions for pathogenic microorganisms such as Salmonella sp. [21, 22]. Garcia et al. and Callaway et al. reported that bifidobacteria can prevent or reduce diseases caused by pathogens, protecting thus consumers’ health [16, 23]. Based on our study results, raffia sap fermented by B. bifidum can be effective against salmonellosis due to Salmonella proliferation.
Figure 5 demonstrates the response surface of L. fermentum and B. bifidum against E. coli. As in the case with Salmonella sp., only B. bifidum showed a significant antagonistic effect on E. coli (P = 0.041, Table 2) at a 10% probability level. An increase in the seeding rate of B. bifidum considerably increased the antagonistic effect, with the maximum inhibition zone of 18 mm.
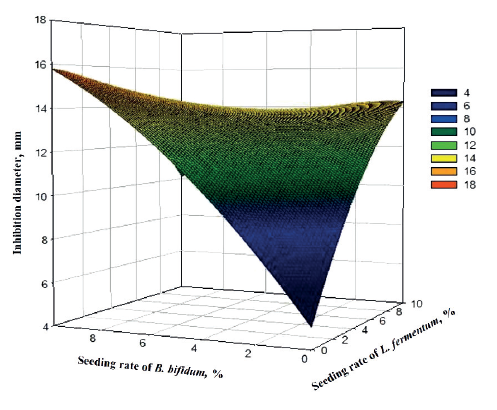
Indeed, lactic acid bacteria exert a strong antagonistic activity against several microorganisms, including those causing the deterioration of food and pathogenic microbes such as E. coli [4, 24]. In addition, the antimicrobial effect of some probiotic extends the shelf life of food [25]. This effect is mainly due to the production of organic acids (lactic acid) and also the production of antimicrobial compounds such as hydrogen peroxide, diacetyl, acetaldehyde, amino acid isomers and bacteriocins [19, 26].
It is important to remember that E. coli is a Gramnegative mammalian intestinal bacterium that makes up about 80% of the aerobic intestinal flora in humans [27, 28]. However, some strains of E. coli can be pathogenic, resulting in gastroenteritis, urinary tract infections, meningitis, or sepsis. Therefore, consumption of raffia sap fermented by B. bifidum can prevent and control the pathogenicity of E. coli.
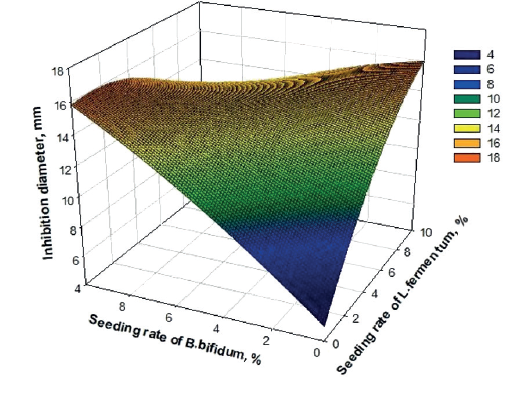
Figure 6 presents the response surface of the mathematical model of inhibition zone of L. fermentum and B. bifidum against B. cereus. Both L. fermentum and B. bifidum individually had a significant antagonistic effect (P = 0.058 and 0.072, respectively, Table 2), whereas their combination was a highly effective (P = 0.007). B. cereus had similar sensitivities to both probiotics in raffia sap (Fig. 6). Inhibition zones reached 18 mm when the seeding rates of L. fermentum and B. bifidum were maximum. The acids and antimicrobial compounds secreted by L. fermentum and B. bifidum in raffia sap are thus a pathway to be exploited to treat diseases, although rare, due to consumption of B. cereus-infected foods. B. cereus is a well-known food-borne pathogen that is ubiquitously distributed in nature and is frequently responsible for food poisoning [20].
Effect of L. fermentum and B. bifidum on L. monocytogenes and optimization of the antagonistic effect. In the case of L. monocytogenes, response surface curves were not required because neither of the probiotic bacteria in raffia sap had a significant antagonistic effect (P = 0.722 for L. fermentum and P = 0.259 for B. bifidum, Table 2). This can be explained by the greater resistance of this bacterium to acidity [29]. Probably, the fermentation time should be increased to enhance the antagonistic properties of the raffia sap drink, but it would make the drink more acidic and hence undrinkable. It would be better to exploit this hypothesis in the context of the synthesis, isolation and production of biologically active compounds from raffia sap fermented by L. fermentum and B. bifidum.
In conclusion, the optimization of the antagonistic effect was done on the basis of specifications that aimed to maximize the inhibition zones. Thus, an optimal antagonistic effect would be given by seeding rates of L. fermentum and B. bifidum of 2 and 10%, respectively, incubation time of 48 h, and temperature of 37°C.
ВЫВОДЫ
The results obtained in this study revealed that raffia sap fermented by probiotics (Lactobacillus fermentum and Bifidobacterium bifidum) had antibacterial properties against bacteria such as Escherichia coli, Listeria monocytogenes, Salmonella sp., and Bacillus cereus which can sometimes be pathogenic. However, further studies should be carry out to determine the mechanism of action of this finding and to confirm its beneficial effect in animal models.КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that there is no conflict of interests related to this article.БЛАГОДАРНОСТИ
We would like to thank Prof. Ali Ahmed and Prof. Nso Emmanuel for the internship research in IUT and ENSAI of Ngaoundere. We also thank the LAGETA (Food Process and Technology laboratory) and LBP (Bioprocess Laboratory) of the University of Ngaoundere for technical assistance.СПИСОК ЛИТЕРАТУРЫ
- Salminen S. Human studies on probiotics: aspects of scientific documentation. Scandinavian Journal of Nutrition/ Naringsforskning. 2001;45(1):8–12.
- Chen H, Tian M, Chen L, Cui X, Meng J, Shu G. Optimization of composite cryoprotectant for freeze-drying Bifidobacterium bifidum BB01 by response surface methodology. Artificial Cells Nanomedicine and Biotechnology. 2019;47(1):1559–1569. https://doi.org/10.1080/21691401.2019.1603157.
- Schrezenmeir J, de Vrese M. Probiotics, prebiotics, and synbiotics – approaching a definition. American Journal of Clinical Nutrition. 2001;73(2):361S–364S.
- Heller KJ. Probiotic bacteria in fermented foods: product characteristics and starter organisms. American Journal of Clinical Nutrition. 2001;73(2):374S–379S.
- Tadesse G, Ephraim E, Ashenafi M. Assessment of the antimicrobial activity of lactic acid bacteria isolated from Borde and Shamita, traditional Ethiopian fermented beverages, on some foodborne pathogens and effect of growth medium on inhibitory activity. Internet Journal of Food Safety. 2005;5:13–20.
- Saarela M, Mogensen G, Fonden R, Matto J, Mattila-Sandholm T. Probiotic bacteria: safety, functional and technological properties. Journal of Biotechnology. 2000;84(3):197–215. https://doi.org/10.1016/s0168-1656(00)00375-8.
- Yu HS, Lee NK, Choi AJ, Choe JS, Bae CH, Paik HD. Antagonistic and antioxidant effect of probiotic Weissella cibaria JW15. Food Science and Biotechnology. 2019;28(3):851–855. https://doi.org/10.1007/s10068-018-0519-6.
- Lim SM, Im DS. Inhibitory effects of antagonistic compounds produced from Lactobacillus brevis MLK27 on adhesion of Listeria monocytogenes KCTC3569 to HT-29 cells. Food Science and Biotechnology. 2012;21(3):775–784. https://doi.org/10.1007/s10068-012-0101-6.
- Saxelin M, Tynkkynen S, Mattila-Sandholm T, de Vos WM. Probiotic and other functional microbes: from markets to mechanisms. Current Opinion in Biotechnology. 2005;16(2):204–211. https://doi.org/10.1016/j.copbio.2005.02.003.
- Ratsep M, Naaber P, Koljalg S, Smidt I, Shkut E, Sepp E. Effect of Lactobacillus plantarum strains on clinical isolates of Clostridium difficile in vitro. Journal of Probiotics and Health. 2014;2(1). https://doi.org/10.4172/2329-8901.1000119.
- Arqués JL, Rodríguez E, Langa S, Landete JM, Medina M. Antimicrobial activity of lactic acid bacteria in dairy products and gut: Effect on pathogens. Biomed Research International. 2015;2015. https://doi.org/10.1155/2015/584183.
- Kasra-Kermanshahi R, Mobarak-Qamsari E. Inhibition effect of lactic acid bacteria against food born pathogen, Listeria monocytogenes. Applied Food Biotechnology. 2015;2(4):11–19.
- Mbarga MJA, Desobgo ZSC, Ngouné TL, Meisam Z, Engeribo A, Maryam B. Producing probiotic beverage based on raffia sap fermented by Lactobacillus fermentum and Bifidobacterium bifidum. Research on Crops. 2019;20(3):629–634. https://doi.org/10.31830/2348-7542.2019.092.
- Sukhikh AS, Zakharova YuV, Yuzhalin AE, Bykov AT, Kotova TV, Poznyakovskiy VM. Criteria for standardization of probiotic components in functional food products. Foods and Raw Materials. 2018;6(2):457–466. https://doi.org/10.21603/2308-4057-2018-2-457-466.
- Ojo OC, Agboola SA. Evaluation of the antagonistic activity of bacteria isolated from palm wine (Raphia vinifera) on Salmonella typhi from different sources. Journal of Advances in Microbiology. 2018;13(1):1–9.
- Garcıa P, Rodrıguez L, Rodrıguez A, Martınez B. Food biopreservation: Promising strategies using bacteriocins, bacteriophages and endolysins. Trends in Food Science and Technology. 2010;21(8):373–382. https://doi.org/10.1016/j.tifs.2010.04.010.
- Devaraj NK, Suppiah S, Veettil SK, Ching SM, Lee, KW, Menon RK, et al. The effects of probiotic supplementation on the incidence of diarrhea in cancer patients receiving radiation therapy: A systematic review with meta-analysis and trial sequential analysis of randomized controlled trials. Nutrients. 2019;11(12). https:/doi.org/10.3390/nu11122886.
- Luquet F-M, Corrieu.G. Bactéries lactiques et probiotiques. Lavoisier; 2008. 320 p.
- Cintas LM, Casaus MP, Herranz C, Nes IF, Hernandez PE. Review: Bacteriocins of lactic acid bacteria. Food Science and Technology International. 2001;7(4):281–305. https://doi.org/10.1106/r8de-p6hu-clxp-5ryt.
- Hwang JY, Park JH. Characteristics of enterotoxin distribution, hemolysis, lecithinase, and starch hydrolysis of Bacillus cereus isolated from infant formulas and ready-to-eat foods. Journal of Dairy Science. 2015;98(3):1652–1660. https://doi.org/10.3168/jds.2014-9042.
- Aslim B, Yuksekdag ZN, Sarikaya E, Beyatli Y. Determination of the bacteriocin-like substances produced by some lactic acid bacteria isolated from Turkish dairy products. LWT – Food Science and Technology. 2005;38(6):691–694. https://doi.org/10.1016/j.lwt.2004.08.001.
- Soleimani NA, Kermanshahi RK, Yakhchali B, Sattari TN. Antagonistic activity of probiotic lactobacilli against Staphylococcus aureus isolated from bovine mastitis. African Journal of Microbiology Research. 2010;4(20):2169–2173.
- Callaway TR, Edrington TS, Anderson RC, Harvey RB, Genovese KJ, Kennedy CN, et al. Probiotics, prebiotics and competitive exclusion for prophylaxis against bacterial disease. Animal Health Research Reviews. 2008;9(2):217–225. https://doi.org/10.1017/S1466252308001540.
- Akarca G, Tomar O, Güney İ, Erdur S, Gök V. Determination of sensitivity of some food pathogens to spice extracts. Journal of Food Science and Technology. 2019;56(12):5253–5261. https://doi.org/10.1007/s13197-019-03994-1.
- O’Sullivan L, Ross RP, Hill C. Potential of bacteriocin-producing lactic acid bacteria for improvements in food safety and quality. Biochimie. 2002;84(5–6):593–604. https://doi.org/10.1016/s0300-9084(02)01457-8.
- Ju J, Xie Y, Guo Y, Cheng Y, Qian H, Yao W. The inhibitory effect of plant essential oils on foodborne pathogenic bacteria in food. Critical Reviews in Food Science and Nutrition. 2019;59(20):3281–3292. https://doi.org/10.1080/10408398.2018.1488159.
- You YS, Cha YJ, Choi SS. Effect of lactic acid bacteria on intestinal E. coli in Caenorhabditis elegans. Food Science and Biotechnology. 2015;24(5):1853–1858. https://doi.org/10.1007/s10068-015-0242-5.
- Abdelhamid AG, El-Masry SS, El-Dougdoug NK. Probiotic Lactobacillus and Bifidobacterium strains possess safety characteristics, antiviral activities and host adherence factors revealed by genome mining. EPMA Journal. 2019;10(4):337–350. https://doi.org/10.1007/s13167-019-00184-z.
- Poroś-Głuchowska J, Markiewicz Z. Antimicrobial resistance of Listeria monocytogenes. Acta Microbiologica Polonica. 2003;52(2):113–129.
- Lee VC. The antibiotic resistance crisis. Part 1: Causes and threats. P&T. 2015;40(4):277–283.
- Unal Turhan E, Erginkaya Z, Uney MH, Ozer EA. Inactivation effect of probiotic biofilms on growth of Listeria monocytogenes. Kafkas Universitesi Veteriner Fakultesi Dergisi. 2017;23(4):541–546. https://doi.org/10.9775/kvfd.2016.17253.
