

Аннотация
Introduction. Gliadin proteins are one of the gluten fractions. They are soluble in alcoholic solution and divided into four groups (α + β, γ, ω1.2, and ω5-gliadins). In this paper gliadins were extracted from wheat flour, and optimal conditions for their separation were determined.Study objects and methods. The separation was performed by capillary gel electrophoresis on Agilent apparatus, CE 7100 (a capillary with an inner diameter of 50 μm, a total length of 33 cm, and an effective length of 23.50 cm). In order to determine the optimal conditions, different solvent concentrations (50, 60, and 70% ethanol), capillary temperatures (20, 25, 30, 35, and 40°C), and electrode voltages (–14.5, –16.5, –17.5 and –18.5 kV) were applied. Migration time and relative concentration of each protein molecules within gliadin fractions in the electrophoregram were analysed using Agilent ChemStation Software.
Results and discussion. The optimal conditions for gliadin separation were: solvent 70% (v/v) ethanol, capillary temperature of 25°C, and electrode voltage of –16.5 kV. Under these conditions, the total proteins were indetified as Xav = 23.50, including α + β gliadin fraction (Xav = 7.50 and relative concentration RC = 28.29%), γ-gliadins (Xav = 5.00, RC = 26.66%), ω1.2-gliadins (Xav = 4.33, RC = 14.93%), and ω5-gliadins (Xav = 6.67, RC = 30.98%).
Conclusion. The results of the research can be of fundamental importance in the study of gluten proteins and the influence of technological procedures on their change and the possibility of reducing the allergic effect of gluten during processing.
Ключевые слова
Proteins, wheat, extraction, ethanol, electrophoresis, glutenВВЕДЕНИЕ
Gliadin proteins represent one of the gluten fractions. Most gliadin proteins are present as monomers. They affect the viscosity and extensibility of wheat flour [1, 2]. Gliadins are divided into four groups, namely α-, β-, γ- and ω-gliadins. This division is based on mobility at low pH, i.e. in acidic conditions of A-PAGE electrophoresis medium (acid polyacrylamide gel electrophoresis). Based on research that was later conducted on amino acid sequences, α and β gliadins were classified in the same group (α/β) [3–5].
Modern methods, such as two-dimensional electrophoresis and high-pressure liquid chromatography with reversed phase, allow the separation of gliadin fractions into more than a hundred components. Based on the analysis of amino acid sequences (complete and partial), amino acid composition and molecular weight, gliadins are divided into: ω5, ω1.2, α + β and γ [3, 6–8]. ω-gliadins are characterized by a high content of glutamine, proline and phenylalanine. These amino acids together make up about 80% of the total ω gliadin composition. ω5-gliadins have a higher molecular weight (≈ 50 000 Da) than ω1.2 (≈ 40 000 Da). Most ω gliadins lack cysteine, so there is no possibility of disulfide binding. These proteins consist of repetitive sequences that are rich in glutamine and proline [3, 9, 10].
Molecular weights of α + β and γ-gliadins overlap (≈ 28 000–35 000 Da). The content of glutamine and proline is much lower compared to ω-gliadin. They differ in tyrosine content. Each of the two types has an N- and a C-terminal region [3, 11]. The N-terminal region (40–50% of total proteins) consists of repeating amino acid sequences that are rich in glutamine, proline, phenylalanine, and tyrosine. The repeating sequences of α + β gliadin are dodecapeptides. They are repeated five times. A typical unit of γ-gliadin is repeated up to 16 times. They are interspersed with additional remains [12, 13]. Within the C-terminal region α + β and γ-gliadins are homologous. The sequences are not repeting. They contain less glutamine and proline than the N-terminal region and have a more common composition. α + β and γ gliadins contain six or eight cysteine residues. These residues are located in the C-terminal region. They form intramolecular disulfide bonds [14, 15]. Although the content of total gliadin proteins depends on the type of wheat and growth conditions (soil, climate, fertilization), α + β and γ-gliadins are the highest components. Ω-gliadins are present in lower amounts [16–18].
To separate gliadin proteins, the following techniques are used: high performance liquid chromatography with reversed phase RP-HPLC, exclusion chromatography SE-HPLC, high performance capillary electrophoresis HPCE, sodium dodecyl sulphate polyacrylamide gel electrophoresis SDS-PAGE, and isoelectric focusing IEF [19]. One of the newer techniques for gliadin separation is the highperformance SDS-GCE, which is based on the difference in electrophoretic mobility of ions in solution within the capillary. The molecule size affects the mobility of ions [20].
The number of people who are allergic to gluten proteins from wheat is increasing, which makes food producers give their consumers a guarantee that products declared as “gluten free” really do not contain gluten. The aim of this study was to investigate optimal conditions (solvent concentrations, capillary temperature and voltage) for their separation by high performance capillary gel electrophoresis.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Gliadin extraction. We analyzed gliadins in wheat flour samples (ash content: max 0.55%, moisture max: 15%, acidity: max 3, protein content 9.8 g/100 g) purchased on the market of the Republic of Srpska, Bosnia and Herzegovina by capillary gel electrophoresis.
Extraction of gliadin proteins was performed according to a modified Osborne method, as described by Lookhart and Bean [21]. After the albumins and globulins were removed (extraction was performed 3 times with 8 mL of deionized water each, it was obtained in laboratory conditions, on the apparatus Siemens water Technologies W3T199551, Siemens Ultra Clear, at a conductivity of 0.055 mS/cm and at a temperature of 20°C and 3 times with 8 mL of 2% solution of NaCl, NaCl, Lach-Ner, Czech Republic, high purity, ≥ 99.00%) gliadin was extracted with 8 mL of ethanol of different concentrations (50, 60 and 70% v/v, refined REAHEM, 96% v/v ethyl alcohol, Srbobran, quality corresponds to the quality property for ethyl alcohol, contains a minimum of 96% v/v ethanol). Samples were homogenized on a vortex (Advanced Vortex Mixer ZX3, 3000 rpm) for 30 min. The samples were then centrifuged in a centrifuge (Rotina 380 R, Hettich Zentrifugen) for 5 min at 1000 rpm. The resulting supernatant was poured into a normal 25 mL vessel, and after the third extraction the normal vessel was made up to final volume with ethanol of various concentrations (50, 60 and 70% v/v). The precipitate was then washed with deionized water.
Samples preparation for analysis at GCE. Prior to analysis samples were diluted with sample buffer (SDS-MW sample buffer, PA 800 plus, Beckman Coulter, USA), so that the total volume was 95 μL and the concentration was 1 mg/mL. Then 2 μL of internal standard (10 kDa, PA 800 plus, Beckman Coulter, United States) and 5 μL of 2-mercaptoethanol (high purity, 99.00%, Sigma-Aldrich Chemie GmbH, Germany) were added. The samples were then heated on a thermo-shaker (Thermo-Shaker, TS-100, Biosan) at 100°C for 3 min. After cooling to room temperature for 5 min, the samples were ready for analysis by capillary gel electrophoresis (Agilent, CE 7100).
Preparation SDS-MW standard for analysis by capillary gel electrophoresis. Prior to the preparation standard, based on the recommendation of the kit manufacturer, the standard was taken to room temperature for 15 min after removal from the refrigerator. It was then carefully stirred on a vortex (Advanced Vortex Mixer ZX3, 3000 rpm) for a few seconds. After that, 10 μL of standard (SDS-MW standard, PA 800 plus, Beckman Coulter, United States) was pipetted into the vial. Then 85 μL of buffer (SDS-MW sample buffer, PA 800 plus, Beckman Coulter, USA) and 2 μL of internal standard (10 kDa, PA 800 plus, Beckman Coulter, USA) were added. Then 5 μL of 2-mercaptoethanol (Sigma-Aldrich Chemie GmbH, Germany, high purity, 99.00%) was added. Then, it was heated on a thermo-shaker (Thermo- Shaker, TS-100, Biosan), at a temperature of 100°C for 3 min. After heating, the standard vial was cooled to room temperature over 5 min. Prepared in this way, the standard is ready for analysis.
Gliadin proteins separation by capillary gel electrophoresis. Separation of gliadin proteins by capillary gel electrophoresis was performed on an Agilent apparatus, CE 7100, with a capillary inner diameter of 50 μm, a total length of 33 cm, and an effective length of 23.50 cm. The SDS-MW analysis kit, PA 800 plus (2015 Beckman Coulter, USA) was used for separation. SDS gel buffer (0.2% SDS, pH = 8) was used to fill the capillary. The kit contains the following chemicals: SDS-MW gel buffer (0.2% SDS, pH = 8), SDS-MW sample buffer (100 mM Tris-HCl, pH = 9, 1% SDS), internal standard (10 kDa), external standard (10 to 225 kDa), acid wash solution (0.1N HCl), base wash solution (0.1N NaOH), as well as two capillaries 57 cm long, 50 μm ID. According to the manufacturer’s instructions, the kit is stored at room temperature after opening, except for the internal and external standards, which are stored at a temperature of 2–6°C. Preparation of the capillary electrophoresis (CE) instrument was done according recommendations Agilent Technologies [22–24].
Statistical data processing. Statistical data processing was performed in IBM SPSS, Statistics 26. Descriptive statistical analysis calculated the average value, standard deviation and 95% confidence interval of the average value. Variance analysis of different groups was used to evaluate the effect of solvent concentrations, capillary temperature and electrode voltage on the number of detected proteins and the relative concentration of each gliadin proteins.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
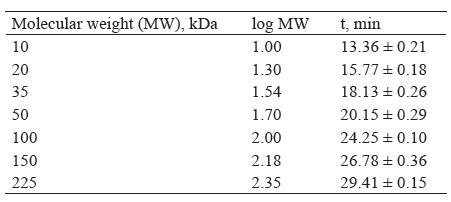
In order to determine molecular weights unknown proteins, a calibration curve was obtained using 7 proteins in SDS-MW size standard.
Electrophoregram, the migration time, and the calibration curve of MW standard proteins with known molecular weight (10, 20, 35, 50, 100, 150 and 225 kDa) are presented in Fig. 1, Table 1, and Fig. 2, respectively.
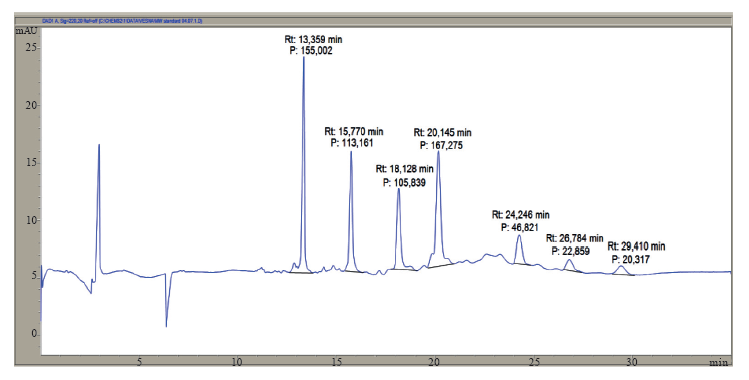

The proteins were separated by capillary gel electrophoresis (CE, Agilent, CE 7100, internal capillary diameter 50 μm, total capillary length 33 cm, effective capillary length 23.50 cm, capillary temperature 25°C, voltage –16.5 kV (reverse mode), duration of analysis 30 min, and absorbance measured at 220 nm).
The ratio of molecular weights (log MW) and migration time (t) of proteins is represented by the equation y = 0.08168x – 0.00098, where y represents logMW and x represents the migration time of proteins (t). R2 shows the correlation coefficient (0.9847).
A calibration curve was used to estimate the molecular weight of unknown proteins. The coefficient of correlation shows a high dependence of the logarithm of the molecular weight of the protein and the migration time of the protein.
The number of proteins in each gliadin fraction and their relative concentration were obtained based on the total number of identified proteins and the total relative concentration.
Table 2 shows descriptive indicators of total proteins and the number of gliadin proteins after extraction with different concentrations of ethanol.
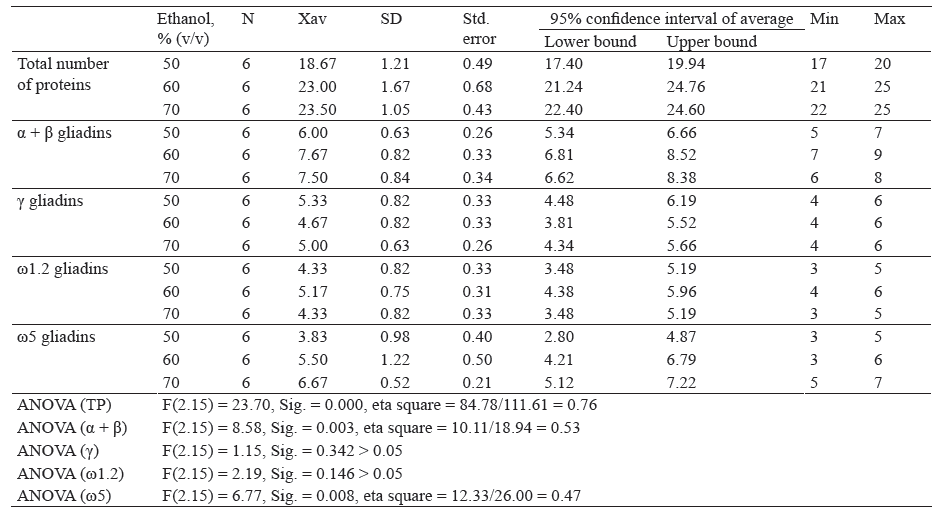
Descriptive analysis showed that the highest number of proteins (23.50) was obtained after extraction with 70% ethanol, by the method of Lookhart and Bean. The lowest number of proteins was obtained by extraction with 50% ethanol (18.67). One-factor analysis of the variance of different groups showed that there was a statistically significant difference in the number of proteins, F(2.15) = 23.70, Sig. = 0.000. The highest number of within α + β gliadin fractions was obtained after extraction with 60% ethanol (7.67). The lowest number of those proteins was obtained after extraction with 50% ethanol (6.00). A statistically significant difference was found in the number of proteins, F(2.15) = 8.58, Sig. = 0.003.
Extraction with 50 and 60% ethanol produced the highest and the lowest number of proteins within the γ-gliadins (5.33 and 4.67, respectively). There was no statistically significant difference in the number of p roteins, F (2.15) = 1.15, Sig. = 0.342. The h ighest amount of ω1.2-gliadins was obtained after extraction with 60% ethanol (5.17), while the lowest after extraction with 50 and 70% ethanol (4.33). One-factor variance analysis showed no statistically significant difference, F(2.15) = 2.19, Sig. = 0.146. T he h ighest n umber of ω5-gliadins was obtained after extraction with 70% ethanol (6.67). The lowest amount was observed after extraction with 50% ethanol (3.83). A statistically significant difference in the number of proteins was found, F(2.15) = 6.77, Sig. = 0.008.
According to Table 2, an increasing ethanol concentration increased total proteins, increased and then slightly decreased α + β gliadin fraction, decreased and then increased γ-gliadins, increased and then decreased ω1.2-gliadins, and increased ω5 gliadin fractions.
Table 3 shows descriptive indicators of the total relative concentration and the relative concentration of gliadin proteins after extraction with different concentrations of ethanol.
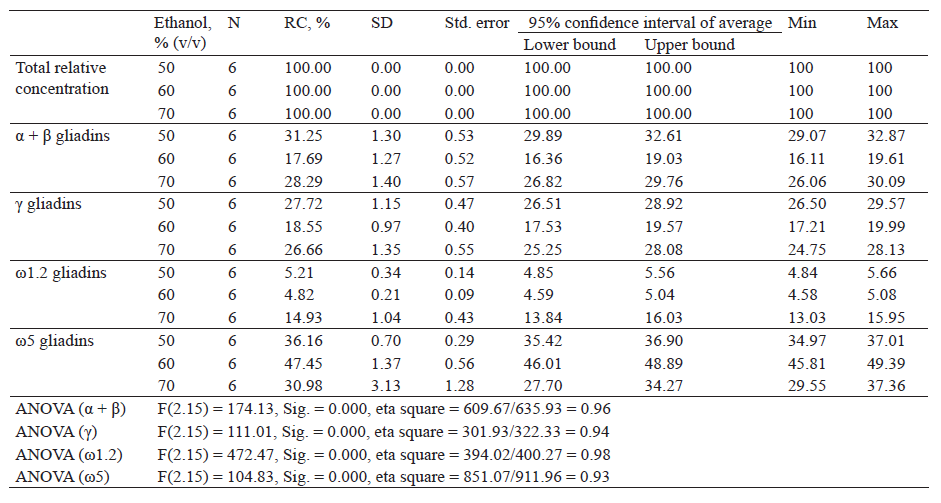
Descriptive analysis showed the highest relative protein concentration of α + β gliadin fractions after extraction with 50% ethanol (31.25%) and the lowest concentration after extraction with 60% ethanol (17.69%). One-factor variance analysis revealed a statistically significant difference in the relative protein concentration, F(2.15) = 174.13, Sig. = 0.000.
Extraction with 50 and with 60% ethanol produced the highest and the lowest relative concentration of γ-gliadins (27.72 and 18.55%). A statistically significant difference in relative concentration was found, F(2.15) = 111.01, Sig. = 0.000. The relative concentration of ω1.2-gliadins was the highest after extraction with 70% ethanol (14.93%) and the lowest after extraction with 60% ethanol (4.82%). The one-factor analysis of the variance showed a statistically significant difference in the relative concentration, F(2.15) = 472.47, Sig. = 0.000.
As for ω5 gliadin fractions, they were found in the highest concentration after extraction with 60% ethanol (47.45%) and the lowest after extraction with 70% ethanol (30.98%). There was a statistically significant difference in the relative concentration, F(2.15) = 104.83, Sig. = 0.000.
Based on the obtained results (Table 3), an increasing ethanol concentration decreased and then increased the relative concentration of α + β, γ- and ω1.2-gliadins and increased and then decreased that of ω5-gliadins.
Table 4 shows descriptive indicators of the total number of proteins and number of gliadin proteins separated by fractions after extraction with 70% (v/v) ethanol and separated at a capillary temperature of 20, 25, 30, 35 and 40°C.
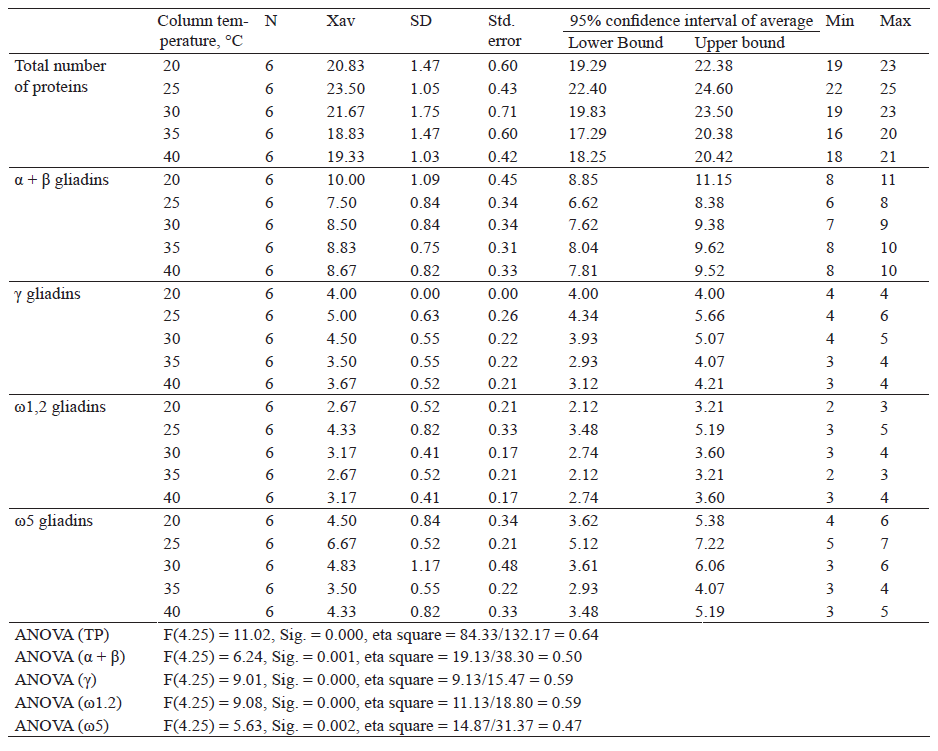
Descriptive analysis revealed that the highest number of proteins was obtained after extraction with 70% ethanol and at a capillary temperature of 25°C (23.50), while the lowest amount of proteins was observed at 35°C (18.83). The one-factor analysis of variance showed a statistically significant difference in the number of proteins, F(4.25) = 11.02, Sig. = 0.000.
The highest and the lowest numbers of α + β gliadin fractions were obtained at 20°C (10.00) and 25°C (7.50), respectively. There was a statistically significant difference, F(4.25) = 6.24, Sig. = 0.001. The number of γ-gliadins was the highest at 25°C (5.00) and the lowest at 35°C (3.50). ANOVA test showed a statistically significant difference in the number of proteins, F(4.25) = 9.01, Sig. = 0.000. The highest amount of ω1.2-gliadins was obtained at a capillary temperature of 25°C (4.33) and the lowest at 20 and 35°C (2.67). A statistically significant difference in the number of proteins was found, F(4.25) = 9.08, Sig. = 0.000. ω5-gliadins were identified in the highest number at a capillary temperature of 25°C (6.67) and in the lowest number at 35°C (3.50). The one-factor analysis of variance revealed a statistically significant difference, F(4.25) = 5.63, Sig. = 0.002.
According to the results obtained, it can be seen that with increasing capillary temperature, total proteins increased, then decreased and increased slightly again. α + β gliadin fractions decreased, then increased and decreased slightly again. As for γ-, ω1.2- and ω5-gliadins, their fractions increased, then decreased and increased slightly again.
Table 5 shows descriptive indicators of the total relative concentration of proteins and relative concentration of gliadin proteins separated by fractions after extraction with 70% (v/v) ethanol and separated at different capillary temperatures.
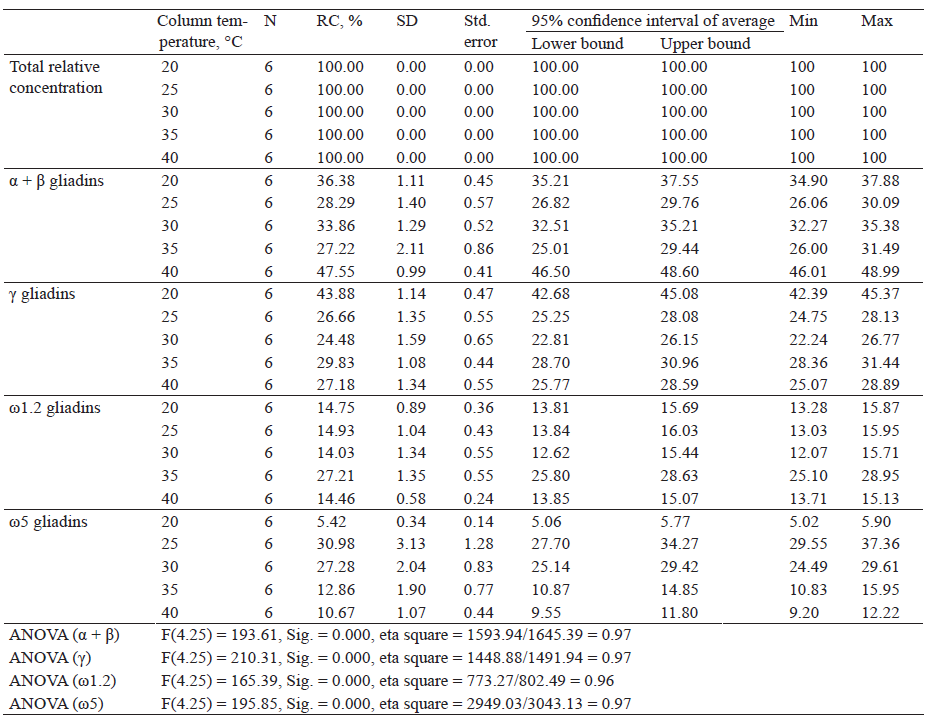
According to the data, the highest relative concentration of α + β gliadin fractions was obtained after extraction with 70% ethanol and a capillary temperature of 40°C (47.55%). The lowest concentration was observed at 35°C (27.22%). One-factor variance analysis revealed a statistically significant difference in the relative concentration, F(4.25) = 193.61, Sig. = 0.000. The relative concentration of γ-gliadins was the highest at 20°C (43.88%) and the lowest at 30°C (24.48%). A statistically significant difference in the relative concentration of different groups was F(4.25) = 210.31, S ig. = 0 .000. A c apillary t emperature of 35°C led to the highest relative concentration within the ω1.2-group (27.21%), while 30°C provided the lowest (14.03%). There was a statistically significant difference in the relative concentration, F(4.25) = 165.39, Sig. = 0 .000. T he h ighest r elative c oncentration o f ω 5- gliadins was obtained after extraction with 70% ethanol and at a capillary temperature of 25°C (30.98%) and the lowest at 20°C (5.42%). The effect of capillary temperature on relative protein concentration within ω5 gliadin fraction was examined by one-factor analysis of variance. A statistically significant difference in the relative concentration within the fraction was found, F(4.25) = 195.85, Sig. = 0.000.
Based on the obtained results (Table 5), it can be seen that with increasing capillary temperature, the relative concentration of α + β gliadins decreased, then increased, decreased, and increased again. Within γ-gliadins, the relative concentration decreased, then increased, and decreased again. The relative concentration of ω1.2-gliadins increased, then decreased, increased again and finally decreased. Within the ω5 gliadin fractions, the relative concentration increased and then decreased.
Table 6 shows descriptive indicators of total proteins and the number of gliadin proteins separated by fractions after extraction with 70% (v/v) ethanol and separated applying different electrode voltages (reverse mode).
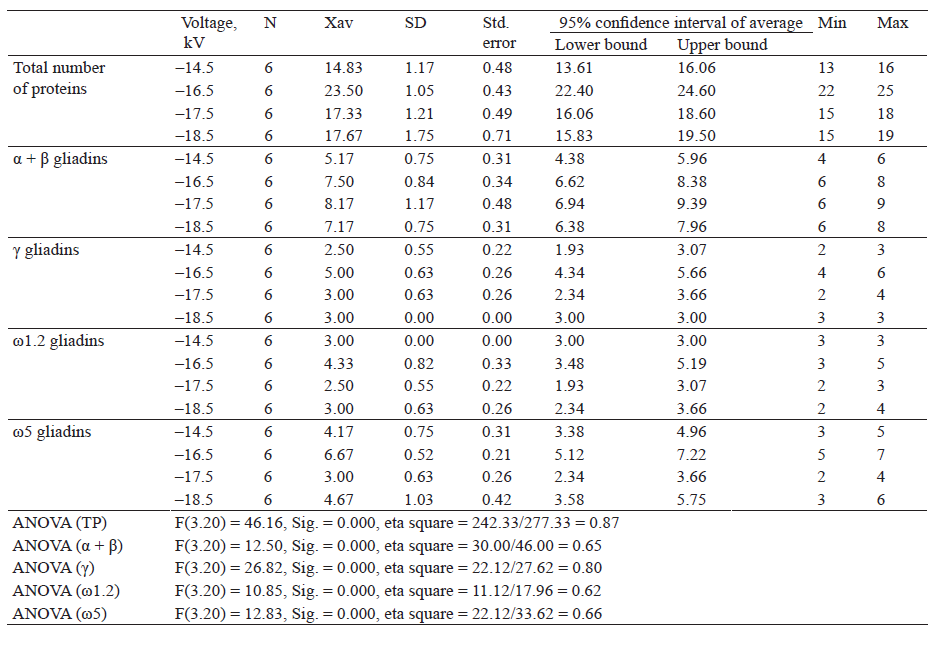
The highest number of proteins was obtained after extraction with 70% ethanol, according to the method by Lookhart and Bean and electrophoretic separation at a voltage of –16.5 kV (23.50). The lowest number of proteins was obtained at –14.5 kV (14.83). It was found that there is a statistically significant difference in the number of proteins, F(3.20) = 4 6.16, Sig. = 0.000. The highest and the lowest amounts of proteins within the α + β gliadin fractions were obtained at a voltage of –17.5 Kv and –14.5 kV (8.17 and 5.17, respectively). A statistically significant difference in the number of proteins was revealed, F(3.20) = 12.50, Sig. = 0.000. Within γ-gliadins, the highest amount of proteins was obtained at a voltage of –16.5 kV (5.00) and the lowest at –14.5 kV (2.50). ANOVA test showed a statistically significant difference in the number of proteins, F(3.20) = 2 6.82, S ig. = 0 .000. T he h ighest n umber o f proteins within the ω1.2 gliadin fractions was obtained at a voltage of –16.5 kV (4.33). The lowest amount of ω1.2 gliadins was obtained at –17.5 kV (2.50). A statistically significant difference in the number of proteins w as f ound, F (3.20) = 1 0.85, S ig. = 0 .000. Descriptive analysis showed that the highest number of proteins within the ω5 gliadin fractions was at –16.5 kV (6.67) and the lowest at –17.5 kV (3.00). There was a statistically significant difference in the number of proteins, F(3.20) = 12.83, Sig. = 0.000.
We can seen that with increasing voltage, total proteins increased, then decreased and increased slightly again. Within the α + β and γ gliadin fractions, the number of proteins increased and then decreased. Within the fraction of ω1.2- and ω5-gliadins, the amount of proteins increased, then decreased and increased slightly again.
Table 7 shows descriptive indicators of the total relative concentration of proteins and relative concentration of gliadin proteins separated by fractions after extraction with 70% (v/v) ethanol and separated by applying different electrode voltages (reverse mode).

Descriptive analysis showed that the highest relative concentration of α + β gliadins was obtained at a voltage of –17.5 kV (65.13%). The lowest concentration within this fraction was at –16.5 kV (28.29%). A statistically significant difference in relative concentration was F(3.20) = 851.47, Sig. = 0.000. The h ighest a nd the lowest relative concentrations of γ-gliadins were obtained at –14.5 kV and at –18.5 kV (27.37 and 21.87%, respectively). A statistically significant difference in the relative protein concentration was found, F(3.20) = 2 0.47, Sig. = 0.000. A voltage of –14.5 kV caused the highest (26.73%) and –18.5 the lowest (3.91%) relative concentration of ω1.2-gliadins. The one-factor analysis of variance showed a statistically significant difference, F(3.20) = 1316.91, Sig. = 0.000. Relative concentration of ω5-gliadins obtained at a voltage of –18.5 kV was the highest (40.30%) and at –17.5 kV the lowest (4.91%). There was a statistically significant difference in the relative concentration, F(3.20) = 549.81, Sig. = 0.000.
According to the results obtained, the increasing voltage decreased then increased and decreased again the relative concentration of α + β gliadin proteins. Within the fraction of γ- and ω1.2- gliadins the concentration decreased, and ω5-gliadins increased then decreased and increased again.
Lookhart and Bean performed separation and characterization of wheat proteins by high-pressure capillary electrophoresis (HPCE) [21]. Gliadins were extracted with 70% (v/v) ethanol. Separation of proteins was performed at a voltage of 22 kV and at a temperature of 45°C. The detection wavelength was 200 nm. Based on the obtained results, the retention time of gliadin proteins was: α gliadins 3–4 min (molecular weight according to SDS-PAGE 35–38 kDa), β 4–6 min (37–43 kDa), γ 5–6 min (43–47 kDa), and ω 6.8–10 min (48–63 kDa).
Bietz and Schmalzried analyzed gliadins from wheat by capillary electrophoresis [25]. Gliadins were extracted with ethanol and methanol of different concentrations (30, 40, 50, 60 and 70% v/v), with and without the reducing agent dithioerythritol. The temperature of the capillary ranged from 30 to 50°C, and the voltage from 8 to 12 kV. The detection wavelength was 200 nm. Capillary temperature of 40°C and voltage 10 kV showed optimal conditions. Ethanol proved to be a better solvent than methanol.
Changing ethanol concentration (50, 60, and 70% v/v), capillary temperature (20, 25, 30, 35, and 40°C), and voltage (–14.5; –16.5, –17.5, and –18.5 kV), we found that the optimal conditions for separation of gliadin proteins were 70% ethanol concentration, a capillary temperature of 25°C, and a voltage of –16.5 kV (reverse mode) (Fig. 3).

Our results are in agreement with Lookhart and Bean and Bietz and Schmalzried [21, 25]. Although the mentioned authors separated gliadin proteins by using different techniques of capillary electrophoresis, 70% ethanol proved to be the optimal solvent, which lines up with our results [21]. The gliadin proteins in this work were separated in less than 10 min, which is in agreement with Lookhart and Bean [21].
ВЫВОДЫ
Based on the results obtained, the optimal conditions for gladin separation were 70% ethanol concentration, a capillary temperature of 25°C, and a voltage of –16.5 kV (reverse mode). Under these conditions, total proteins were 23.5, including α + β gliadin proteins (Xav = 7.50, relative concentration 28.29%), γ-fractions (Xav = 5.00, RC = 26.66%), ω1.2-gliadins (Xav = 4.33, RC = 14.93%), and ω5-gliadins (Xav = 6.67, RC = 30.98%).
The results obtained in this paper can greatly contribute to the prevention of the incorrect declaration of the “gluten free” products, reduction of health risks for people who are sensitive to gluten proteins, as well as the cost of treating the ones with celiac disease.
In addition, the results of the research are of fundamental importance in the study of gluten proteins and the influence of technological procedures on their change and the possibility of reducing the allergic effect of individuals gluten proteins, during processing.
КОНФЛИКТ ИНТЕРЕСОВ
The authors declare no potential conflict of interest.ФИНАНСИРОВАНИЕ
This work is a part of the research project “Analysis gluten proteins holders of allergic reactions by high performance liquid chromatography”, number 19/6-020/96123/18, financed by the Ministry of Scientific and Technological Development, Higher Education and Information Society Republic of Srpska.СПИСОК ЛИТЕРАТУРЫ
- Shan L, Khosla C. Chemistry and biology of gluten proteins. Immunology, Endocrine and Metabolic Agents in Medicinal Chemistry. 2007;7(3):187–193. DOI: https://doi.org/10.2174/187152207780832397.
- Urade R, Sato N, Sugiyama M. Gliadins from wheat grain: an overview, from primary structure to nanostructures of aggregates. Biophysical Reviews. 2018;10(2):435–443. DOI: https://doi.org/10.1007/s12551-017-0367-2
- Wieser H. Chemistry of gluten proteins. Food Microbiology. 2007;24(2):115–119. DOI: https://doi.org/10.1016/j.fm.2006.07.004.
- Zhang Y, Luo G, Liu D, Wang D, Yang W, Sun J, et al. Genome-, transcriptome- and proteome wide analyses of the gliadin gene families in Triticum urartu. PloS ONE. 2015;10(7). DOI: https://doi.org/10.1371/journal.pone.0131559.
- Balakireva AV, Zamyatnin AA. Properties of gluten intolerance: Gluten structure, evolution, pathogenicity and detoxification capabilities. Nutrients. 2016;8(10). DOI: https://doi.org/10.3390/nu8100644.
- Wieser H. Relation between gliadin structure and coeliac toxicity. Acta Paediatrica. 1996;85(412):3–9. DOI: https://doi.org/10.1111/j.1651-2227.1996.tb14239.x.
- Qian Y, Preston K, Krokhin O, Mellish J, Ens W. Characterization of wheat gluten proteins by HPLC and MALDI TOF mass spectrometry. Journal of the American Society for Mass Spectrometry. 2008;19(10):1542–1550. DOI: https://doi.org/10.1016/j.jasms.2008.06.008.
- Lexhaller B, Colgrave ML, Scherf KA. Characterization and relative quantitation of wheat, rye, and barley gluten protein types by liquid chromatography-tandem mass spectrometry. Frontiers in Plant Science. 2019;10. DOI: https://doi.org/10.3389/fpls.2019.01530.
- Seilmeier W, Valdez I, Mendez E, Wieser H. Comparative investigations of gluten proteins from different wheat species II. Characterization of ω-gliadins. European Food Research and Technology. 2001;212(3):355–363. DOI: https://doi.org/10.1007/s002170000260.
- Horváth C. Storage proteins in wheat (Triticum aestivum L.) and the ecological impacts affecting their quality and quantity, with a focus on nitrogen supply. Journal of Agricultural and Environmental Sciences. 2014;1(2):57–76.
- Cebolla Á, Moreno ML, Coto L, Sousa C. Gluten immunogenic peptides as standard for the evaluation of potential harmful prolamin content in food and human specimen. Nutrients. 2018;10(12). DOI: https://doi.org/10.3390/nu10121927.
- Helmerhorst EJ, Zamakhchari M, Schuppan D, Oppenheim FG. Discovery of a novel and rich source of glutendegrading microbial enzymes in the oral cavity. PloS ONE. 2010;5(10). DOI: https://doi.org/10.1371/journal.pone.0013264.
- Shewry P. What is gluten – Why is it special? Frontiers in Nutrition. 2019;6. DOI: https://doi.org/10.3389/fnut.2019.00101.
- Grosch W, Wieser H. Redox reactions in wheat dough as affected by ascorbic acid. Journal of Cereal Science. 1999;29(1):1–16. DOI: https://doi.org/10.1006/jcrs.1998.0218.
- Srinivasan B, Focke-Tejkl M, Weber M, Pahr S, Baar A, Atreya R, et al. Usefulness of recombinant γ-gliadin 1 for identifying patients with celiac disease and monitoring adherence to a gluten-free diet. The Journal of Allergy and Clinical Immunology. 2015;136(6):1607–1618. DOI: https://doi.org/10.1016/j.jaci.2015.04.040.
- Daniel C, Triboi E. Effects of temperature and nitrogen nutrition on the grain composition of winter wheat: Effects on gliadin content and composition. Journal of Cereal Science. 2000;32(1):45–56. DOI: https://doi.org/10.1006/jcrs.2000.0313.
- Wieser H, Kieffer R. Correlations of the amount of gluten protein types to the technological properties of wheat flours determined on a micro-scale. Journal of Cereal Science. 2001;34(1):19–27. DOI: https://doi.org/10.1006/jcrs.2000.0385.
- Hurkman WJ, Tanaka CK, Vensel WH, Thilmony R, Altenbach SB. Comparative proteomic analysis of the effect of temperature and fertilizer on gliadin and glutenin accumulation in the developing endosperm and flour from Triticum aestivum L. cv. Butte 86. Proteome Science. 2013;11(1). DOI: https://doi.org/10.1186/1477-5956-11-8.
- Malvano F, Albanese D, Pilloton R, Di Matteo M. A new label-free impedimetric aptasensor for gluten detection. Food Control. 2017;79:200–206. DOI: https://doi.org/10.1016/j.foodcont.2017.03.033.
- Sirén H. Capillary electrophoresis in food analysis. In: Nollet LML, Toldra F, editors. Handbook of food analysis – Two Volume Set. Boca Raton: CRC Press; 2015. pp. 493–519. DOI: https://doi.org/10.1201/b18668.
- Lookhart G, Bean S. Separation and characterization of wheat protein fractions by high-performance capillary electrophoresis. Cereal Chemistry. 1995;72(6):527–532.
- Wenz C. Performance of commercially available gels for protein characterization by capillary gel electrophoresis with UV detection on the Agilent 7100 CE System. Application Note [Internet]. [cited 2020 Jul 1]. Available from: https://www.gimitec.com/file/5990-7976EN.pdf.
- Braud C, Devarieux R, Atlan A, Ducos C, Vert M. Capillary zone electrophoresis in normal or reverse polarity separation modes for the analysis of hydroxy acid oligomers in neutral phosphate buffer. Journal of hromatography B: Biomedical Sciences and Applications. 1998;706(1):73–82. DOI: https://doi.org/10.1016/s0378-4347(97)00468-4.
- Grujić R, Savanović D. Analysis of myofibrillar and sarcoplasmic proteins in pork meat by capillary gel electrophoresis. Foods and Raw Materials. 2018;6(2):421–428. DOI: https://doi.org/10.21603/2308-4057-2018-2-421-428.
- Bietz JA, Schmalzried E. Capillary electrophoresis of wheat gliadin: initial studies and application to varietal identification. LWT – Food Science and Technology. 1995;28(2):174–184. DOI: https://doi.org/10.1016/s0023-6438(95)91346-7.
