

Аннотация
Introduction. Increased resistance of microorganisms to traditional antibiotics has created a practical need for isolating and synthesizing new antibiotics. We aimed to study the antibiotic activity and resistance of bacteriocins produced by lactic acid bacteria and other microorganisms.Study objects and methods. We studied the isolates of the following microorganism strains: Bacillus subtilis, Penicillium glabrum, Penicillium lagena, Pseudomonas koreenis, Penicillium ochrochloron, Leuconostoc lactis, Lactobacillus plantarum, Leuconostoc mesenteroides, Pediococcus acidilactici, Leuconostoc mesenteroides, Pediococcus pentosaceus, Lactobacillus casei, Lactobacillus fermentum, Bacteroides hypermegas, Bacteroides ruminicola, Pediococcus damnosus, Bacteroides paurosaccharolyticus, Halobacillus profundi, Geobacillus stearothermophilus, and Bacillus caldotenax. Pathogenic test strains included Escherichia coli, Salmonella enterica, Staphylococcus aureus, Pseudomonas aeruginosa, Bacillus mycoides, Alcaligenes faecalis, and Proteus vulgaris. The titer of microorganisms was determined by optical density measurements at 595 nm.
Results and discussion. We found that eleven microorganisms out of twenty showed high antimicrobial activity against all test strains of pathogenic and opportunistic microorganisms. All the Bacteroides strains exhibited little antimicrobial activity against Gramnegative test strains, while Halobacillus profundi had an inhibitory effect on Gram-positive species only. The Penicillium strains also displayed a slight antimicrobial effect on pathogenic test strains.
Conclusion. The antibiotic resistance of the studied lactic acid bacteria and other bacteriocin-producing microorganisms allows for their use in the production of pharmaceutical antibiotic drugs.
Ключевые слова
Lactic acid bacteria, bacteriocins, antibiotic properties, antibiotic resistance, natural sources, isolatesВВЕДЕНИЕ
New microorganisms that are resistant to traditional antibiotics have recently become known to medicine [1]. Today, large numbers of people worldwide are dying from various infections caused by antibiotic-resistant strains of microorganisms [2]. Therefore, there is an increasingly important scientific and practical need for new antimicrobial drugs with a wide spectrum of action.
Modern researchers are actively studying bacteriocins produced by Gram-positive bacteria, which are antibiotic proteins [3]. Due to their complex structure, bacteriocins can be classified as peptides with different activity, gene control, and biochemical processes [4, 5]. They do not develop antimicrobial resistance and therefore are widely used in medicine and pharmacology [4, 6]. These substances are known for high antibiotic activity against closely related strains of microorganisms. Lactic acid microorganisms are among the most effective producers of bacteriocins and bacteriocin-like agents [3].
Bacteriocinogenesis has apparently evolved as a result of adaptation and survival in a harmful environment, having occupied a certain niche in microbiology [7]. Bacteriocins are produced by lactic acid bacteria – natural microbiota in the digestive system of humans and animals, as well as in food raw materials, products, or animal feed. Bacteriocins colonize natural and industrial substrates [8–10]. Most often, they do not dominate over saprophytic microorganisms of spore and non-spore forms, over cocci, yeasts, molds, and Gram-negative bacteria, which inhibit antibiotics [11, 12].
Bacteriocin production is a complex process that requires optimal parameters to affect the system. Not all bacteria can synthesize bacteriocins. It has been proved that the ability to synthesize a small amount of bacteriocinogenic substances by individual strains is hereditary [13, 14]. However, the synthesis can be improved by genetic engineering, DNA-tropic substances ultraviolet rays, peroxides, chemical mutagens, and other agents [15, 16]. Since mid-20th century, extensive experiments have been in operation to create new bacteriocin-producing bacteria.
A number of Gram-positive strains, such as Lactobacillus, Streptococcus, Bacillus, Mycobacterium, Staphylococcus, Corynebacterium, Leuconostoc, Sarcina, Micrococcus, Clostridium and Streptomyces, have been reported to synthesize bacteriocins [2, 3, 17, 18].
A lot of current research is focused on bacteriocins produced by lactic acid microorganisms. For example, diacetin B-1, a bacteriocin isolated from Lactococcus lactis, consists of 37 amino acid residues and has a molecular weight of 4300 Da [19–21]. Scientists know of 14 strains of Lactococcus lactis capable of synthesizing bacteriocins. All bacteriocins inhibit the growth of S. aureus, P. acidilactici, L. Plantarum, and many Listeria species [14, 22, 23].
Amylovorin 471, a bacteriocin produced by Lactobacillus amylovorus DCE 471, is used as a biopreservative in food and feed [24].
A purified form of enterocin A obtained from Enterococcus faecium contains 47 amino acid residues, including 4 cystine residues, and has a molecular weight of 4289 Da. Enterocin A has a similar amino acid sequence to that of nisin, a bacteriocin produced by lactic acid bacteria [25].
Bacteriocins are also formed by other types of enterococci. For example, E. faecalis S-48 produces a 80 kDa bacteriocin that is sensitive to proteases and has an inhibitory effect on E. faecalis [26].
Thus, many infectious diseases can be prevented and treated by isolating new strains of lactic acid microorganisms that produce bacteriocins with antibacterial action [27, 28]. Unlike Lactobacillus strains, the antimicrobial activity of Lactococcus strains has not been well studied [2, 14].
Therefore, there is an urgent need for isolating new antimicrobial and antibiotic-resistant bacteriocins formed by lactic acid bacteria and other antagonist microorganisms, as well as studying their properties and prospects for the pharmaceutical industry [29, 30].
We aimed to study the antibiotic activity and resistance of bacteriocins produced by lactic acid bacteria and other antagonist microorganisms isolated from natural systems in the Kemerovo region.
In particular, we aimed to:
– study the antimicrobial effect of lactic acid
bacteria and other antagonist bacteriocin-producing
microorganisms on pathogenic and opportunistic
microflora that can cause severe infectious diseases in
humans;
– select the isolates of microorganisms with bacteriocin
properties (antimicrobial activity) to determine their
antibiotic resistance; and
– examine the resistance of lactic acid bacteria and other
antagonist microorganisms to the main antibiotics of
various series.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Microbial communities in various habitats (soil, water, animal gastrointestinal tract, animal products, etc.) were used as natural systems from which we isolated strains of bacteriocin-producing microorganisms. The sampling took place in the Kemerovo region.
Our objects of study included the isolates of bacteriocin-producing microorganism strains, such as Bacillus subtilis, Penicillium glabrum, Penicillium lagena, Pseudomonas koreenis, Penicillium ochrochloron, Leuconostoc lactis, Lactobacillus plantarum, Leuconostoc mesenteroides, Pediococcus acidilactici, Leuconostoc mesenteroides, Pediococcus pentosaceus, Lactobacillus casei, Lactobacillus fermentum, Bacteroides hypermegas, Bacteroides ruminicola, Pediococcus damnosus, Bacteroides paurosaccharolyticus, Halobacillus profundi, Geobacillus stearothermophilus, and Bacillus caldotenax.
Prior to isolation, we incubated microorganisms on an agar medium melted and poured into Petri dishes (covering a third or a quarter of the area), then sterilized and cooled. The incubation lasted 4–5 days at 30°C (until complete or almost complete sporulation by vegetative cells). Then, the grown colonies were suspended in 30 mL of a sterile liquid T3 medium. The flasks with the inoculated medium were placed on an orbital shaker (220 rpm, 72–80 h, 30°C). The stage of sporulation was determined by phase contrast microscopy. At the end of incubation, we found 98–100% of spores and crystals in the liquid medium in relation to the vegetative cells. The number of colony-forming units per mL of culture fluid (CFU/mL) was determined with a series of dilutions followed by incubation in Petri dishes with a T3 medium (five replicates) for 24 h at 30°C. After incubation, we counted the grown colonies and expressed the results in CFU/ml, or spores/ml in our case.
We used the following pathogenic test strains:
– Escherichia coli ATCC 25922 – opportunistic bacteria
causing gastroenteritis in humans;
– Salmonella enterica ATCC 14028 – pathogenic
bacteria causing gastroenteritis in humans;
– Staphylococcus aureus ATCC 25923 – pathogenic
bacteria causing pneumonia, meningitis, osteomyelitis,
endocarditis, infectious toxic shock and sepsis in
humans;
– Pseudomonas aeruginosa B6643 – opportunistic
bacteria causing nosocomial infections in humans;
– Bacillus mycoides EMTC 9 (Russian collection of
extremophilic microorganisms and type cultures) – opportunistic
bacteria causing foodborne toxic infections
in humans;
– Alcaligenes faecalis EMTC 1882 – opportunistic
bacteria causing septicemia and meningitis in newborns
and intra-abdominal infections in adults;
– Proteus vulgaris ATCC 63 – opportunistic bacteria
causing acute intestinal infections in humans.
Cultivation of microorganism test strains. Escherichia coli ATCC 25922 was cultivated on a medium composed of 10 g tryptone, 5 g yeast extract, 10 g sodium chloride, and 1 L water (pH 7.5–8.0, 37°C). Salmonella enterica ATCC 14028 was cultivated on a medium composed of 10 g peptic digest of animal tissue, 5 g meat extract, 5 g glucose, 4 g sodium hydrogen phosphate, 0.3 g iron sulfate, 8 g bismuth sulfite, 0.025 g brilliant green, 20 g agar-agar, and 1 L water (pH 7.5–7.9, 35°С).
Staphylococcus aureus ATCC 25923 was cultivated on a medium composed of 10 g casein hydrolysate, 2.5 g yeast extract, 30 g gelatin, 10 g D-mannitol, 55 g sodium chloride, 75 g ammonium sulfate, 5 g potassium hydrogen phosphate, 15 g agar-agar, and 1 L water (рН 6.8–7.2, 30°С).
Pseudomonas aeruginosa B6643 was cultivated on a medium composed of 1 L meat water, 5 g NaCl, and 10 g peptone (рН 6.8–7.0, 37°С).
Bacillus mycoides EMTC 9 was cultivated on a medium composed of 10 g casein hydrolysate, 2.5 g yeast extract, 5 g glucose, 2.5 g potassium hydrogen phosphate, 3 g agar-agar, and 1 L water (рН 7.2–7.6, 30°С).
Alcaligenes faecalis EMTC 1882 was cultivated on a medium composed of 10 g special peptone, 5 g sodium chloride, 0.3 g sodium azide, 0.06 g chromogenic mixture, 2 g Tween-80, 1.25 g sodium hydrogen phosphate, 15 g agar-agar, and 1 L water (рН 7.3–7.5, 37°С).
Proteus vulgaris ATCC 63 was cultivated on a medium composed of 8 g peptone, 5 g sodium chloride, 1 g sodium deoxycholate, 1.5 g chromogenic mixture, 10.5 g propylene glycol, 15 g agar-agar, and 1 L water (рН 7.1–7.5, 37°С).
The quantity of microorganisms (titer) in the suspensions of overnight broth cultures grown on standard media was determined by optical density measurements at 595 nm.
Lactic acid bacteria and other antagonist microorganisms isolated from natural sources in the Kemerovo region were assessed for their antimicrobial action in two ways, using the diffusion method and measuring optical density.
Diffusion method. Test strain bacteria inoculated onto an agar medium using the spread plate technique were immediately covered with paper disks impregnated with the metabolites of microorganisms under study (10 μL/disk). A disc with a nutrient medium was used as a control, and a disc with ciprofloxacin (a standard antibiotic) was used as a reference drug. The plates were incubated for 24 h at a temperature optimal for each test strain. The quantity of microorganisms was determined by measuring the size (mm) of a transparent zone around the disc, indicating the absence of microbial growth [31].
Optical density measurement. Test strain bacteria were incubated with the metabolites in 96-well culture plates [32]. We resuspended broth cultures aged for 12 h in a medium corresponding to the species of microorganisms to inoculate, bringing their amount to ~ 105 CFU/mL. At the same time, we added the cell suspension and the metabolites under study to the wells in an amount of 1/10 of the total volume. A liquid nutrient medium was used as a control and ciprofloxacin was used as a reference drug (10 μg/mL). The total volume of the suspension in the well was 200 μL. The experiments were performed in duplicate. Incubation was carried out on a shaker at 580 rpm at a temperature optimal for each test strain. After 24 h, we measured the optical density on a PICO01 spectrophotometer (Picodrop Limited, UK) at 595 nm. The bactericidal activity was determined by changes in the optical density compared to the control. In the wells where cell growth stopped or slowed down, the optical density was lower than in those with normal growth.
Microbial spores were stained according to the Schaeffer-Fulton method. The method uses a combined effect of a concentrated brilliant green solution and temperature on the impermeable spore membrane with further decolorization of the cytoplasm of a vegetative cell and its contrast staining with safranin. Microscopic examination showed that the spores were stained green and the cells, red. To establish the presence of flagella, we studied the mobility of cultures in the “squashed straw” preparations [33].
The antibiotic resistance was determined by the zones of growth inhibition for the isolates with antibiotic discs. For this, we inoculated isolate cells onto a temporary medium using the spread plate technique, with antibiotic discs on the agar. The experimental results were recorded after 24 h of cultivation in the incubator at 28°C [31].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Table 1 shows the results of using the diffusion method to assess the antimicrobial properties of lactic acid bacteria and other microorganisms isolated from natural sources in the Kemerovo region.
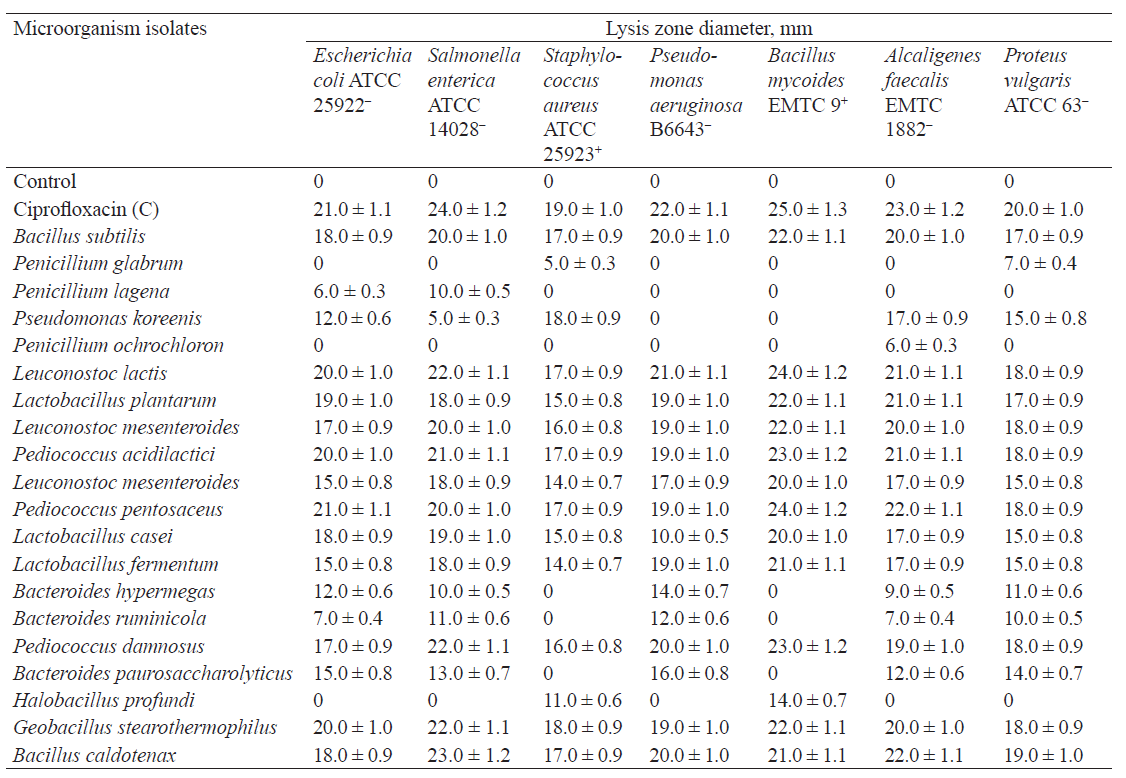
Of the twenty microorganism strains under study, eleven exhibited high antimicrobial activity against all test strains of pathogenic and opportunistic microorganisms (Bacillus subtilis, Leuconostoc lactis, Lactobacillus plantarum, Leuconostoc mesenteroides, Pediococcus acidilactici, Pediococcus pentosaceus, Lactobacillus casei, Lactobacillus fermentum, Pediococcus damnosus, Geobacillus stearothermophilus, Bacillus caldotenax).
Bacteroides hypermegas, Bacteroides ruminicola, and Bacteroides paurosaccharolyticus showed insignificant antimicrobial activity against Gramnegative test strains, while Halobacillus profundi had an inhibitory effect on Gram-positive species only. Penicillium glabrum had a slight antimicrobial effect on Staphylococcus aureus, Proteus vulgaris, and Shigella flexneri; Penicillium lagena, on the test strains of Escherichia coli, Salmonella enterica, Shigella flexneri, Aspergillus flavus, and Penicillium citrinum; Penicillium ochrochloron, on the test strains of Alcaligenes faecalis and Listeria monocytogenes.
For further studies of antibiotic resistance, we selected four isolates with maximum antimicrobial activity against pathogenic and opportunistic test strains, namely Bacillus subtilis, Leuconostoc lactis, Lactobacillus plantarum, and Leuconostoc mesenteroides.
These isolates were tested for antibiotic resistance, i.e. resistance of a strain to one or more antibacterial drugs, or decreased sensitivity (immunity) of a culture to the action of an antibacterial substance.
Antibiotic resistance can develop as a result of natural selection through random mutations and/ or antibiotic exposure. Microorganisms are able to transmit genetic information about antibiotic resistance through horizontal gene transfer. In addition, antibiotic resistance can be induced artificially by genetic transformation, for example, by introducing artificial genes into the genome of a microorganism [13].
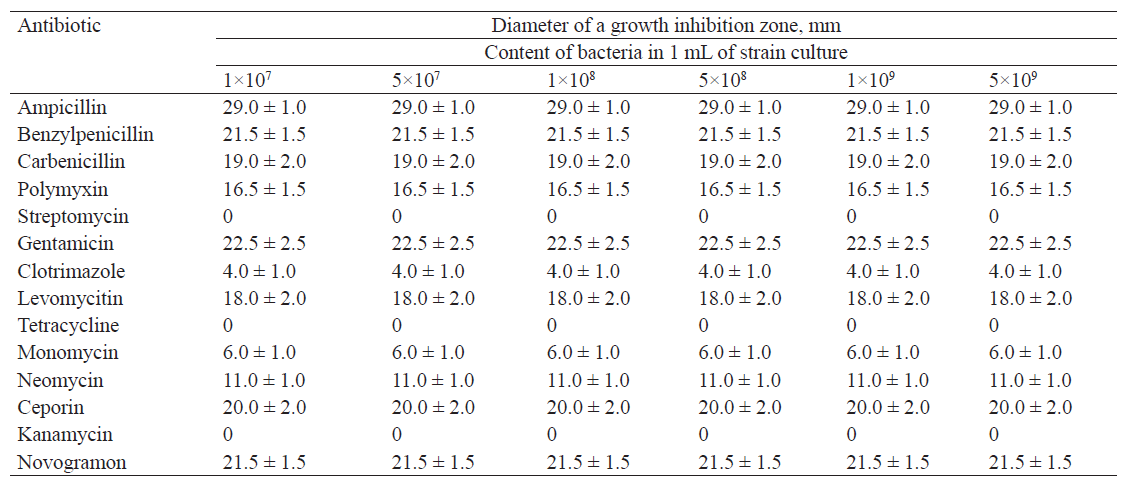
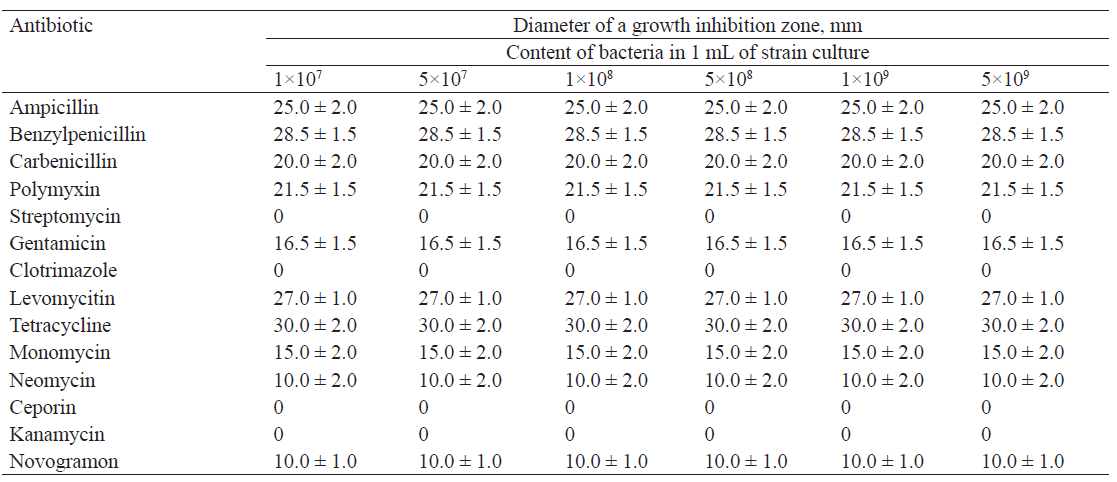
Tables 2–5 show the results of studying the antibiotic resistance of microorganisms isolated from natural sources in the Kemerovo region.
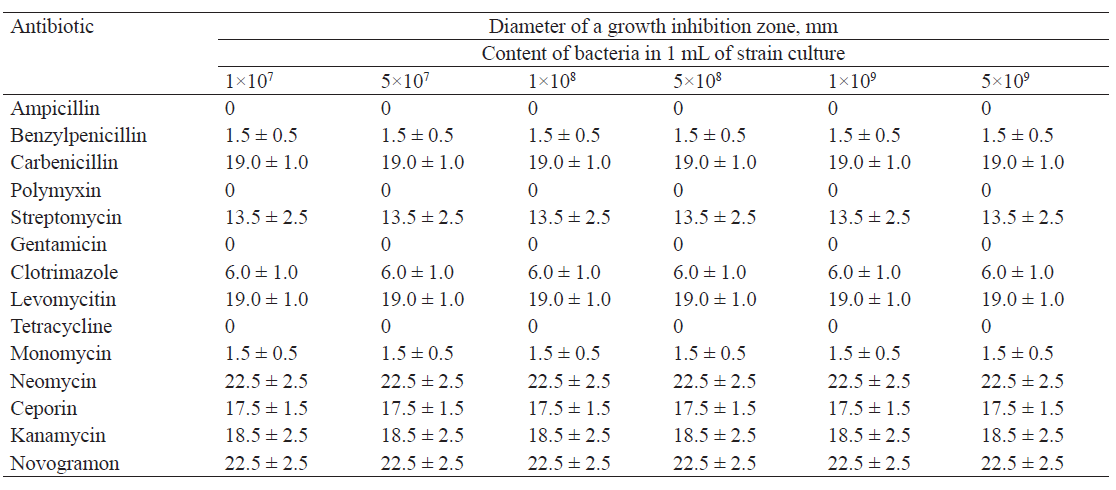

As we can see, Bacillus subtilis proved to be resistant to ampicillin, gentamicin, and tetracycline. It exhibited high sensitivity to neomycin, novogramon, kanamycin, carbenicillin, levomycitin, and ceporin, but low sensitivity to benzylpenicillin, monomycin and clotrimazole.
Leuconostoc lactis was highly sensitive to ampicillin, gentamicin, benzylpenicillin, and novogramon, insensitive to clotrimazole and monomycin, and resistant to streptomycin, tetracycline and kanamycin.
Lactobacillus plantarum showed resistance to streptomycin, clotrimazole, ceporin and kanamycin, high sensitivity to tetracycline, benzylpenicillin, and levomycitin, and low sensitivity to neomycin and novogramon.
Leuconostoc mesenteroides was resistant to streptomycin, tetracycline, and kanamycin, insensitive to clotrimazole and monomycin, and highly sensitive to ampicillin, ceporin, benzylpenicillin, gentamicin, levomycitin, and novogramon.
We found that the isolates with different concentrations of microorganisms displayed the same antibiotic resistance. The diameter of the growth inhibition zone was the same for all concentrations of microorganisms.
ВЫВОДЫ
Thus, we studied the antibiotic activity and resistance of lactic acid bacteria and other antagonist microorganisms isolated from natural sources in the Kemerovo region. We established a correlation between the type of isolate and the type of antibiotic. According to the study, eleven microorganisms out of twenty exhibited high antimicrobial activity, while the rest of the strains had an insignificant effect on the test strains and opportunistic microorganisms.
We found that all the isolates showed some degree of resistance to the following antibiotics used to treat infectious diseases: ampicillin, benzylpenicillin, carbenicillin, polymyxin, streptomycin, gentamicin, clotrimazole, levomycitin, tetracycline, monomycin, neomycin, ceporin, kanamycin, and novogramon.
The progressive resistance of the studied bacteriocinproducing microorganisms to antibiotics allows for their use in the production of pharmaceutical antibiotic drugs.
КОНФЛИКТ ИНТЕРЕСОВ
The authors state that there is no conflict of interest.ФИНАНСИРОВАНИЕ
The study was part of the federal targeted program on “Obtaining Pharmaceutical Substances Based on Antagonist Microorganisms Isolated from Natural Sources” (Unique Project Identifier RFMEFI57418X0207) under Agreements No. 075-02-2018-1934 of December 20, 2018 and No. 075-15-2019-1383 of June 18, 2019 of the Ministry of Science and Higher Education of the Russian Federation (Minobrnauka).СПИСОК ЛИТЕРАТУРЫ
- Cavera VL, Arthur TD, Kashtanov D, Chikindas ML. Bacteriocins and their position in the next wave of conventional antibiotics. International Journal of Antimicrobial Agents. 2015;46(5):494–501. DOI: https://doi.org/10.1016/j.ijantimicag.2015.07.011.
- Bindiya ES, Bhat SG. Marine bacteriocins: A review. Journal of Bacteriology and Mycology: Open Access. 2016;2(5):140–147. DOI: https://doi.org/10.15406/jbmoa.2016.02.00040.
- Yongkiettrakul S, Maneerat K, Arechanajan B, Malila Y, Srimanote P, Gottschalk M, et al. Antimicrobial susceptibility of Streptococcus suis isolated from diseased pigs, asymptomatic pigs, and human patients in Thailand. BMC Veterinary Research. 2019;15(1). DOI: https://doi.org/10.1186/s12917-018-1732-5.
- De Freire Bastos MC, Coelho MLV, da Silva Santos OC. Resistance to bacteriocins produced by Gram-positive bacteria. Microbiology. 2015;161(4):683–700. DOI: https://doi.org/10.1099/mic.0.082289-0.
- Noda M, Miyauchi R, Danshiitsoodol N, Matoba Y, Kumagai T, Sugiyama M. Expression of genes involved in bacteriocin production and self-resistance in Lactobacillus brevis 174A is mediated by two regulatory proteins. Applied and Environmental Microbiology. 2018;84(7). DOI: https://doi.org/10.1128/AEM.02707-17.
- Kumariya R, Garsa AK, Rajput YS, Sood SK, Akhtar N, Patel S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microbial Pathogenesis. 2019;128:171–177. DOI: https://doi.org/10.1016/j.micpath.2019.01.002.
- Ahmad V, Khan MS, Jamal QMS, Alzohairy MA, Al Karaawi MA, Siddiqui MU. Antimicrobial potential of bacteriocins: in therapy, agriculture and food preservation. International Journal of Antimicrobial Agents. 2017;49(1):1–11. DOI: https://doi.org/10.1016/j.ijantimicag.2016.08.016.
- Kassaa I, Hober D, Hamze M, Chihib NE, Drider D. Antiviral potential of lactic acid bacteria and their bacteriocins. Probiotics and Antimicrobial Proteins. 2014;6(3–4):177–185. DOI: https://doi.org/10.1007/s12602-014-9162-6.
- Ghazaryan L, Tonoyan L, Ashhab AA, Soares MIM, Gillor O. The role of stress in colicin regulation. Archives of Microbiology. 2014;196(11):753–764. DOI: https://doi.org/10.1007/s00203-014-1017-8.
- Cramer WA, Sharma O, Zakharov SD. On mechanisms of colicin import: the outer membrane quandary. Biochemical Journal. 2018;475(23):3903–3915. DOI: https://doi.org/10.1042/BCJ20180477.
- Ghequire MGK, De Mot R. The tailocin tale: peeling off phage tails. Trends in Microbiology. 2015;23(10):587–590. DOI: https://doi.org/10.1016/j.tim.2015.07.011.
- Gupta VG, Pandey A. New and future developments in microbial biotechnology and bioengineering. Microbial Secondary Metabolites Biochemistry and Applications. Netherlands: Elsevier; 2019. 213 p.
- Zhao Z, Orfe LH, Liu J, Lu S-Y, Besser TE, Call DR. Microcin PDI regulation and proteolytic cleavage are unique among known microcins. Scientific Reports. 2017;7. DOI: https://doi.org/10.1038/srep42529.
- Ge J, Kang J, Ping W. Effect of acetic acid on bacteriocin production by Gram-positive bacteria. Journal of Microbiology and Biotechnology. 2019;29(9):1341–1348. DOI: https://doi.org/10.4014/jmb.1905.05060.
- Rebuffat S. Microcins and other bacteriocins: bridging the gaps between killing stategies, ecology and applications. In: Dorit RL, Roy SM, Riley MA, editors. The bacteriocins: current knowledge and future prospects. Wymondham: Caister Academic Press; 2016. pp. 11–34. DOI: https://doi.org/10.21775/9781910190371.02.
- Wencewicz TA, Miller MJ. Sideromycins as pathogen-targeted antibiotics. In: Fisher JF, Mobashery S, Miller MJ, editors. Antibacterials. Volume 2. Cham: Springer; 2017. pp. 151–183. DOI: https://doi.org/10.1007/7355_2017_19.
- Garcia-Gutierrez E, O’Connor PM, Colquhoun IJ, Vior NM, Rodriguez JM, Mayer MJ, et al. Production of multiple bacteriocins, including the novel bacteriocin gassericin M, by Lactobacillus gasseri LM19, a strain isolated from human milk. Applied Microbiology and Biotechnology. 2020;104(9):3869–3884. DOI: https://doi.org/10.1007/s00253-020-10493-3.
- Egan K. Ross RP, Hill C. Bacteriocins: antibiotics in the age of the microbiome. Emerging Topics in Life Sciences. 2017;1(1):55–63. DOI: https://doi.org/10.1042/ETLS20160015.
- Alvarez-Sieiro P, Montalbán-López M, Mu DD, Kuipers OP. Bacteriocins of lactic acid bacteria: extending the family. Applied Microbiology and Biotechnology. 2016;100(7):2939–2951. DOI: https://doi.org/10.1007/s00253-016-7343-9.
- Sun Z, Wang X, Zhang X, Wu H, Zou Y, Li P, et al. Class III bacteriocin Helveticin-M causes sublethal damage on target cells through impairment of cell wall and membrane. Journal of Industrial Microbiology and Biotechnology. 2018;45(3):213–227. DOI: https://doi.org/10.1007/s10295-018-2008-6.
- Tracanna V, De Jong A, Medema MH, Kuipers OP. Mining prokaryotes for antimicrobial compounds: from diversity to function. FEMS Microbiology Reviews. 2017;41(3):417–429. DOI: https://doi.org/10.1093/femsre/fux014.
- Acedo JZ, Chiorean S, Vederas JC, van Belkum MJ. The expanding structural variety among bacteriocins from Grampositive bacteria. FEMS Microbiology Reviews. 2018;42(6):805–828. DOI: https://doi.org/10.1093/femsre/fuy033.
- Ongey EL, Yassi H, Pflugmacher S, Neubauer P. Pharmacological and pharmacokinetic properties of lanthipeptides undergoing clinical studies. Biotechnology Letters. 2017;39(4):473–482. DOI: https://doi.org/10.1007/s10529-016-2279-9.
- Wiebach V, Mainz A, Siegert MAJ, Jungmann NA, Lesquame G, Tirat S, et al. The anti-staphylococcal lipolanthines are ribosomally synthesized lipopeptides. Nature Chemical Biology. 2018;14(7):652–654. DOI: https://doi.org/10.1038/s41589-018-0068-6.
- Bennallack PR, Griffitts JS. Elucidating and engineering thiopeptide biosynthesis. World Journal of Microbiology and Biotechnology. 2017;33(6). DOI: https://doi.org/10.1007/s11274-017-2283-9.
- Lajis AFB. Biomanufacturing process for the production of bacteriocins from Bacillaceae family. Bioresources and Bioprocessing. 2020;7(1). DOI: https://doi.org/10.1186/s40643-020-0295-z.
- Crone WJK, Vior NM, Santos-Aberturas J, Schmitz LG, Leeper FJ, Truman AW. Dissecting bottromycin biosynthesis using comparative untargeted metabolomics. Angewandte Chemie-International Edition. 2016;55(33):9639–9643. DOI: https://doi.org/10.1002/anie.201604304.
- Hegemann JD, Zimmermann M, Xie X, Marahiel MA. Lasso peptides: an intriguing class of bacterial natural products. Accounts of Chemical Research. 2015;48(7):1909–1919. DOI: https://doi.org/10.1021/acs.accounts.5b00156.
- Li Y, Ducasse R, Zirah S, Blond A, Goulard C, Lescop E, et al. Characterization of sviceucin from Streptomyces provides insight into enzyme exchangeability and disulfide bond formation in lasso peptides. ACS Chemical Biology. 2015;10(11):2641–2649. DOI: https://doi.org/10.1021/acschembio.5b00584.
- Lear S, Munshi T, Hudson AS, Hatton C, Clardy J, Mosely JA, et al. Total chemical synthesis of lassomycin and lassomycin-amide. Organic and Biomolecular Chemistry. 2016;14(19):4534–4541. DOI: https://doi.org/10.1039/c6ob00631k.
- Garvey M, Rowan NJ. Pulsed UV as a potential surface sanitizer in food production processes to ensure consumer safety. Current Opinion in Food Science. 2019;26:65–70. DOI: https://doi.org/10.1016/j.cofs.2019.03.003.
- Metelev M, Tietz JI, Melby JO, Blair PM, Zhu LY, Livnat I, et al. Structure, bioactivity, and resistance mechanism of streptomonomicin, an unusual lasso peptide from an understudied halophilic actinomycete. Chemistry and Biology. 2015;22(2):241–250. DOI: https://doi.org/10.1016/j.chembiol.2014.11.017.
- Chiorean S, Vederas JC, van Belkum MJ. Identification and heterologous expression of the sec-dependent bacteriocin faerocin MK from Enterococcus faecium M3K31. Probiotics and Antimicrobial Proteins. 2018;10(2):142–147. DOI: https://doi.org/10.1007/s12602-017-9374-7.
- Sukhikh SA, Krumlikov VYu, Evsukova AO, Asyakina LK. Formation and study of symbiotic consortium of lactobacilli to receive a direct application starter. Foods and Raw Materials. 2017;5(1):51–62. DOI: https://doi.org/10.21179/2308-4057-2017-1-51-62.
