

Аннотация
Introduction. In view of the ongoing research into the negative effects of fruit juice on human health, we aimed to study the subchronic toxicity of apple juice, a model mixture based on its components, and ethanol on biomass growth, cellular oxidative enzymes, and chromosomal abnormalities in Allium cepa roots.Study objects and methods. Our objects of study included clarified apple juice and its components such as fructose, glucose, sucrose, D-sorbitol, and malic acid. After treating Allium cepa roots with apple juice and a model mixture in different concentrations, we analyzed their toxic effects on biomass growth, malondialdehyde levels, as well as the nature and frequency of proliferative and cytogenetic disorders in the plant tissues.
Results and discussion. The incubation in an aqueous solution of apple juice at a concentration of 1:5 inhibited the growth in root mass by 50% compared to the control (water). The mitotic index of cells decreased with higher concentrations of juice, reaching zero at a 1:5 dilution. The fructose and model solutions in the same concentrations appeared less toxic in relation to cell mitosis and root mass growth. Although malondialdehyde levels increased in the onion roots treated with juice and model solutions, they were twice as low as in the control due to the juice’s antioxidant activity. Adding 1% ethanol to the 1:2 diluted juice abolished the effect of acute toxicity on root growth and reduced malondialdehyde levels by 30%.
Conclusion. The study revealed a complex of interdependent biomarkers of apple juice responsible for its subchronic toxicity in Allium cepa roots. These data can be used to create biological response models based on the approaches of systems biology and bioinformatics.
Ключевые слова
Juice, fructose, Allium cepa, biotesting, toxicity, cytogenetic analysis, biomarkerВВЕДЕНИЕ
New approaches to food testing are becoming increasingly urgent today, in view of continuously growing production and consumption of various foods. These approaches are primarily meant to ensure food safety by identifying possible toxic effects that food products and related additives may have on human health [1–4]. Any food component can have a negative effect on the human body. Excessive consumption can lead to the accumulation of toxic metabolic products. Some components can cause allergic reactions and modulate adaptation reactions [5]. Such studies are primarily based on in silico and in vivo methods of testing various types of food products.Fruit juice is an integral part of the human diet and, undoubtedly, a complex food system. It contains physiologically active substances (vitamins, minerals, antioxidants, enzymes, and amino acids) that regulate a variety of metabolic processes and increase the body’s resistance to infections. In addition, epidemiological studies have proved that fruits and vegetables reduce the risk of chronic diseases [6, 7]. Clinical studies also confirm that fruit juice can have beneficial effects on blood parameters, cholesterol, and heart function, as well as prevent cancer and Alzheimer’s disease [8–11].
However, the benefits of fruit juice are not as evident as they may seem [12]. As we know, natural mutagens, such as pyrolysidine alkaloids and some flavonoids, account for about 1% of dry matter in almost all higher plants. Moreover, vitamins C, E, and A can have mutagen-potentiating effects [13]. Recent studies have shown that fruits and juices can contribute to the development of cancer and asthma in children [7, 14–16].
Sugars contained in fruit juice and their potentially adverse metabolic effects have long been in the center of scientific debates. Fructose, in particular, is one of the main carbohydrate components of fruit juice. As early as the 1980s, it was considered responsible for several metabolic abnormalities [17, 18]. This carbohydrate can be “toxic”, especially when consumed with sweetened drinks. Moreover, it can participate in the pathogenesis of noncommunicable diseases such as obesity, diabetes, or arthritis [19, 20]. Sucrose, another carbohydrate component of fruit juice, has also shown negative mutagenic effects [21]. In the USA and Europe, a half of sugar consumption accounts for sweetened products with a thick consistency (yogurt, candy and chocolate bars, ice cream, etc.) and the other half, for sweetened fizzy drinks and fruit juices. The negative health effects of fructose have encouraged European countries to impose taxes on sweetened drinks [22].
Quality control is an equally important aspect of fruit juice safety. The past decades have seen a significant increase in the demand for juice, partly due to continuous improvement of its sensory (color, smell, texture, and taste) and technological (convenient packaging, long shelf life) characteristics. As a result, juice composition has undergone a number of changes, with added microelements and synthetic substances (acidity regulators, stabilizers, thickeners, and sweeteners). The technology of juice production (e.g., heat treatment) also affects juice properties. Although the use of these additives is strictly regulated, scientists are increasingly emphasizing a need for rigorous research into the mechanisms of their toxic manifestations [23, 24].
Studies have shown that food additives can lead to cancer and change the functioning of various organs [25–27]. Children are especially vulnerable to their toxic effects that can provoke allergies and other diseases if manufacturers do not follow strict regulations [28]. Although several types of food additives can be used in juice in various combinations, there have been no studies into their integrated toxic effect on the human body. Moreover, as chemically active agents, these additives or their oxidation products can interact with natural organic or inorganic juice compounds and cause especially dangerous mutagenic and carcinogenic effects [29].
In this regard, in vivo studies of subchronic toxicity of fruit juice components are becoming increasingly urgent. Modern food scientists aim to develop models in which the processes of detoxification and metabolism of toxic compounds are similar to those in the human body. At the same time, they strive not to use laboratory animals [4].
We find biotesting quite effective when using plants, in particular Allium cepa roots (Allium test). This test has been successfully used to study toxicity, cytotoxicity, and genotoxicity of various agents, including food additives, as well as to determine genotoxic effects of medicinal plant extracts [23, 30–32]. The Allium test is simple, economical, well reproducible, highly sensitive, applicable in a wide pH range (3.5–11.0), and just as efficient as other biotests. We believe that this test can be reliably used to assess subchronic toxic effects of various juice components, both individually and in combination with each other. Similar studies in animals may not produce objective results. The components under study may be present in the animals’ basic diet, compromising the results.
Our aim was a comparative study of subchronic toxic effects that apple juice, its components, and ethanol have on biomass growth, oxidative enzyme activity at the cellular level, as well as the nature and frequency of proliferative and cytogenetic disorders in Allium cepa roots.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
To model the composition of apple juice, we used the following materials: glucose (SIGMA-ALDRICH, lot. SLBZ9363, Germany), fructose (SIGMA-ALDRICH, lot. SLCC1647, Germany), sucrose (SIGMA-ALDRICH, lot. BCCB2955, Germany), D-sorbitol (SIGMA-ALDRICH, lot.BCBT4918, Germany), malic acid (SIGMAALDRICH, lot.MKBS7851, Germany), and clarified apple juice (10.5% carbohydrates) purchased from a retail outlet.
For biotesting, we used small 5–7 g Allium cepa L. onions of Stuttgart variety with a diameter of 2.5–3 cm, with their dry scaly outer layers removed. The roots were preliminarily germinated in 15 cm3 test tubes with bottled water in a thermostat (23–25°C) for two or three days in complete darkness. The bulbs with a sprouted root length of at least 1 cm were selected for further experiments. Prior to treating them with juice solutions and other compounds listed above, we measured the average root mass in the control group.
Then, the control samples were incubated in water, while the test samples were incubated in aqueous solutions in a thermostat (23–25°С) in complete darkness for 1, 2, or 3 days, depending on the purpose of the experiment. After incubation, the roots were cut off, wiped with filter paper, and weighed [33]. EC50 was determined as a concentration of material that reduced the test function (growth in root mass) by 50% compared to the control, taking into account the average mass of the roots before treatment (except when they were treated with fructose).
For cytogenetic analysis, the cells of the root apical meristem were stained with a 2% aceto-orcein solution (1 g of orcein diluted in 50 cm3 of 45% CH3COOH). For long-term storage, the roots were placed in a 70% ethanol solution used as a preservative. Instant squash preparations were obtained to analyze the division of apical meristem cells, using an Axioskop 40 light microscope (Zeiss). In particular, we determined the mitotic index (ratio of dividing cells to total cells) and the chromosome aberration index (number of chromosomal aberrations related to total cells).
The intensity of lipid peroxidation in root tissues was determined based on the amount of malondialdehyde (MDA) interacting with 2-thiobarbituric acid and expressed in μmol/g (MDA in fresh mass) [34]. We placed 0.2–0.9 (± 0.0001) g into a 15 cm3 polymer tube, added 1 cm3 of trichloroacetic acid (Merck, Germany) at a concentration of 200 g/dm3 and then another 3 cm3 of the same solution after stirring the mixture. The tubes were centrifuged at 1000 g and 4°C for 15 min. Then, we transferred 1 cm3 of the upper liquid layer into another tube and added 4 cm3 of thiobarbituric solution ‒ 0.5 g thiobarbituric acid (Diaem, Russia) and 100 cm3 of trichloroacetic acid (200 g/dm3). The tubes were tightly closed and placed for 30 min in a water bath at 95°C, followed by cooling in an ice bath. Next, the tubes were centrifuged for 10 min at 1000 g and 20°C. The solutions were spectrophotometrically detected on a Cary WinUV 100 spectrophotometer (Varian, USA) at 600 and 532 nm.
Statistical processing was performed in Microsoft Excel and Statistica (version 12). The Student’s criterion and Fisher transformation were used for comparative analysis of percentages.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
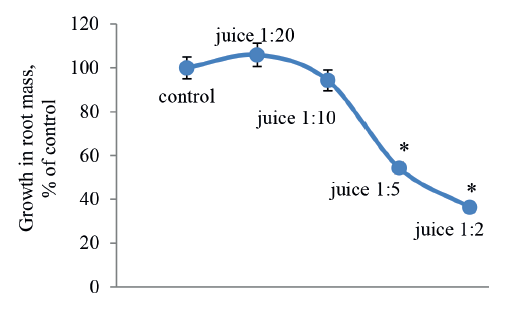
After a three-day sprouting, the onion roots were treated with apple juice diluted with water for 2 days to determine the degree of juice dilution that causes subchronic toxicity. According to the Allium test, toxicity was determined by the changes in root mass after exposure to juice solutions, compared to the control. As we can see in Fig. 1, a decrease in root mass was observed at ten times dilution and EC50 was recorded at five times dilution (P ≤ 0.15).
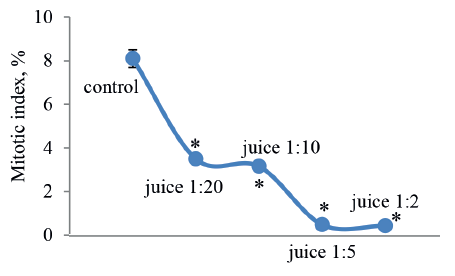
The cytological analysis of the root meristem cells showed that higher juice concentrations decreased the mitotic index more intensively (Fig. 2) than the growth in root mass (Fig. 1). As we can see, the level of proliferation for meristematic cells, when treated with a 1:20 diluted solution of apple juice, was half the control values, and their division almost stopped in the roots with a 50% delay in mass growth (EC50).
As we know, plants grow due to two main processes, cell division and extension. Like all eukaryotes, plant cells enter the cell cycle in response to external mitogenic stimuli. This process is regulated by a large number of compounds, such as phytohormones, ARGOS proteins, CLE peptides, transcription factors, cyclins, and cyclin-dependent protein kinases. Decreased cell proliferation during stress or after treatment with abscisic acid may result from activated expression of genes that encode protein inhibitors of cyclin-dependent protein kinases, ICK/KRP. However, the mechanisms that control differentiation can function independently of the cell cycle [35]. It appears that the subchronic amounts of apple juice triggered similar processes in our study and, therefore, the inhibition of cell proliferation did not significantly affect the growth in root mass.
The percentage of chromosomal aberrations in dividing cells in relation to total stained cells was quite low, about 0.4%, both in the control and the test samples treated with 1:20 and 1:10 diluted juice. We found no effect dependent on the amount. Neither could we determine this indicator in the test samples treated with a higher concentration of juice (1:5 and 1:2 dilution) due to the absence of dividing cells.
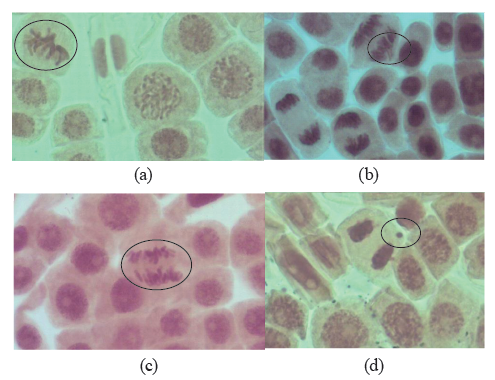
The abnormalities detected in both the control and the test samples included the adhesion of chromosomes to each other, their leading during anaphase, as well as disorganization and disordered separation during metaphase and anaphase. However, these abnormalities were not distributed evenly among the samples. For example, aberrations (Fig. 3) and anaphase leading were almost ten times as high in the test samples. Also, micronuclei were detected during telophase and interphase in the samples treated with tenfold diluted juice.
With the data at hand, we had to understand which of the juice components was responsible for the identified toxic effects and to what extent. Carbohydrates are a major component of apple juice, with up to 10% of fructose, glucose, and sucrose (in 100 g juice). Taking into account published data on the negative effects of glucose on plant growth and development, we conducted several experiments to determine their toxicity for onion roots [36, 37]. We started with fructose, as its content in apple juice is two times as high as that of glucose and sucrose.
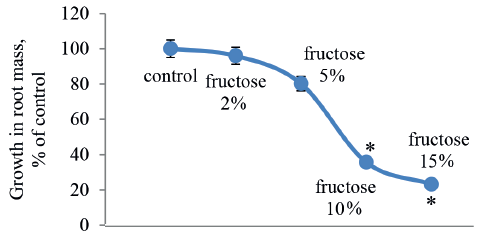
As we can see in Fig. 4, higher concentrations of fructose delayed the growth in root mass, but only a 10% concentration of this carbohydrate revealed a significant difference. After treatment with 10 and 15% fructose solutions, the roots died, becoming thin, soft, and slightly mucous. In the Allium test, this finding probably indicated acute toxicity of fructose in the given concentrations. Thus, the concentration of fructose should not exceed 10%.
Fructose at concentrations of 2 and 5% decreased the mitotic index in the test samples by only 17 and 33%, respectively, compared to the control (Fig. 5). Like in the previous test, the comparative cytogenetic analysis did not reveal a significant increase in the number of chromosomal aberrations, compared to the control. However, we observed some redistribution in their spectrum. For example, higher concentrations of fructose in the test samples caused more disorders such as chromosomal bridges, fragmentation, and segregation (up to 20%), compared to the control.
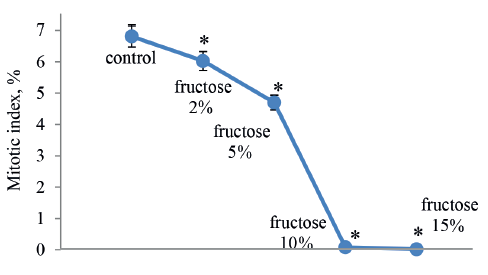
According to the results, the subchronic toxicity of fructose, one of the main components of apple juice, is mainly associated with a weak mitosuppressive effect in the root meristem cells.
Then, we prepared a model aqueous solution from the main chemical components of apple juice. Their concentration ratios corresponded to those in juice [38]. In particular, 100 mL of the model solution contained 7 g fructose, 2 g glucose, 1 g sucrose, 0.5 g D-sorbitol, and 0.3 g malic acid. Prior to that, we had measured the pH of the study objects to make sure that its range was acceptable for the Allium test (Table 1). Next, we analyzed the subchronic toxicity of the resulting model solution and apple juice in Allium cepa roots after two days of germination and two days of treatment.

According to the results, the growth in root mass after treatment with juice was 40% lower than after using the model solution (Table 2), despite the same degree of dilution (1:5, P < 0.05). The cytological analysis showed that the mitotic index of the root meristem cells after treatment with the 1:10 and 1:5 model solutions did not differ much from the control. However, treatment with the 1:10 and 1:5 diluted juice, just like in the previous experiment (Fig. 2), reduced the mitotic index ten times and led to an almost complete halt in cell division. Thus, the chemical components of the model solution, which make up the bulk of juice solids, were not responsible for the subchronic toxicity associated with violation of mitosis in the roots. Obviously, this effect was caused by other juice compounds with antiproliferative activity of natural origin.
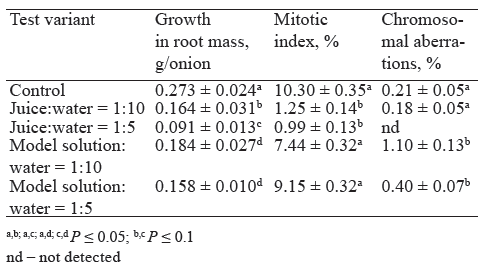
Based on the data, we can conclude that these compounds (one or more) are present in juice in small quantities and have high biological activity. We need further studies to identify these substances and better understand the mechanisms of potential juice toxicity.
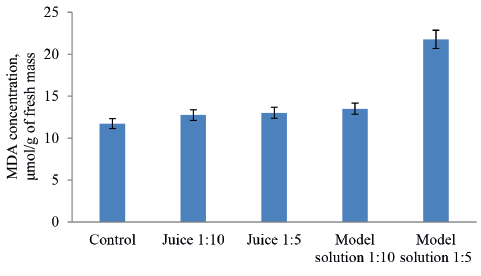
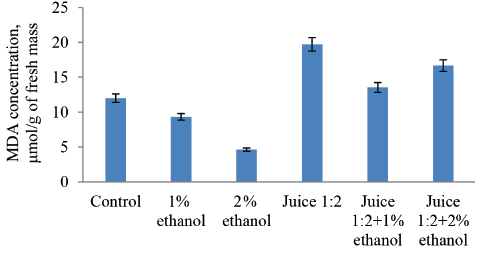
MDA is known to reflect the degree of lipid peroxidation resulting from the oxidation process. The higher its concentration, the more damaged are lipids in the walls of plant cells. In our study, the treatment of onion roots with the 1:10 and 1:5 diluted model solutions produced a dose-dependent increase in MDA, with its maximum levels twice as high as in the control samples (Fig. 6). However, apple juice in the dilutions of 1:10 and 1:5 increased this indicator by only 11%. Apparently, these results are indicative of the juice’s antioxidant activity.
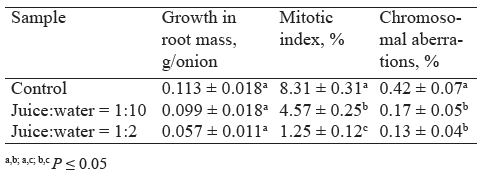
To study toxic effects, we treated the onion roots, which had germinated for two days, with the 1:10 and 1:2 diluted juice for only one day. As we can see in Table 3, a day of incubation brought about a slightly higher (10%) decrease in mitotic indices in these test samples than in those treated for two days, compared to the control (Fig. 2). Thus, the toxic effect was recorded as early as after the first cycle of cell division, while the decrease in root mass growth was more likely to be cumulative.
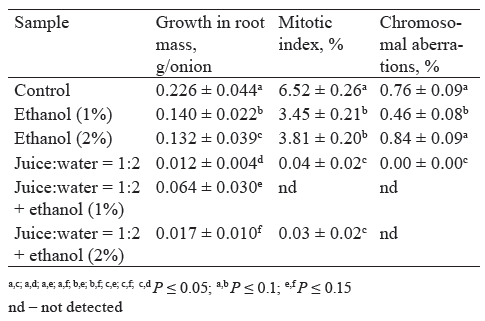
Food additives are commonly studied for toxicity separately from those food products which they are part of. We believe that such practice does not allow scientists to objectively determine the patterns of toxic manifestations. Therefore, our further experiments attempted to evaluate the effect of ethanol on the previously detected toxicity of Allium cepa roots, which had been germinated for two days and then incubated with apple juice for another two days. We chose this food additive due to the fact that 1 and 2% aqueous ethanol solutions delay the growth of Allium cepa roots within EC50 [29, 32]. In addition, ethanol may be part of some juice-containing products.
Table 4 shows that 1% ethanol increased the average mass of the roots treated with the 1:2 diluted juice by a factor of five. We believe that this effect can be associated with the activity of lipid oxidation enzymes and its regulation. Indeed, raising ethanol concentration to 2% not only decreased their activity, but made it lower than the control values (Fig. 4). However, in the 1:2 diluted juice samples, MDA was almost 1.7 times as high as in the control (Fig. 7), which we had expected from the previous results (Fig. 3). Thus, 1% ethanol appeared to slow down the destruction of cell wall lipids caused by the juice components, which had a positive effect on the root growth. The question is, why is it that a higher concentration of ethanol (2%) did not cause a similar effect? Probably, despite lower lipid oxidation, the total toxicity of 2% ethanol was so high that it prevented the roots from growing and developing. Another observation we made was that adding 1 and 2% ethanol to the juice did not increase the proliferative activity of the meristem cells. This result was quite predictable since treating roots with ethanol solutions decreased the mitotic index of the meristem cells, compared to the control.
ВЫВОДЫ
Our study showed that apple juice manifested subchronic toxicity when it was diluted with water in a ratio of 1:5 (~ 2% soluble solids). The toxicity caused a 50% delay in the growth of Allium cepa roots, compared to the control. At the same time, it sharply inhibited the division of meristem cells, with their mitotic index decreasing by a factor of 18 and the MDA concentration increasing by 11%. To identify the mechanisms of these disorders, we treated the roots with the main compounds of juice dry solids – fructose, glucose, sucrose, D-sorbitol, and malic acid – and compared the above indicators. We found that in contrast to the 1:5 diluted juice, 2% fructose decreased the mitotic index by only 17%, compared to the control. The model solution containing 1.4% fructose, 0.4% glucose, 0.2% sucrose, 0.1% D-sorbitol, and 0.06% malic acid showed a 40% higher growth in root mass compared to the 1:5 diluted juice (P < 0.05), the same mitotic index of meristem cells as the control, and a doubled concentration of MDA compared to the control.
Thus, the subchronic toxicity of apple juice primarily manifested through its antiproliferative activity in the meristem cells. However, the above juice components were not involved in that activity. What they were responsible for was an increased level of lipid oxidation in the root tissues, which was restrained by the natural antioxidants present in the juice.
In addition, we analyzed the contribution of a food additive (ethanol) to the potential toxicity of apple juice, using the Allium test. We found that 1% ethanol in the 1:2 diluted juice reduced the concentration of MDA in the roots by 30%, with no effect of acute toxicity in relation to their growth.
The above effects of, and relationships between, various biomarkers of apple juice and its components can form a basis for more detailed large-scale research into its safety. Our findings can also be used to study the toxic potential of juice depending on manufacturing technology or food additives, as well as to create new juice-based products.
КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that they have no conflict of interest.ФИНАНСИРОВАНИЕ
The manuscript was part of a state assignment fulfilled for the V.M. Gorbatov Federal Scientific Center for Food Systems RAS.СПИСОК ЛИТЕРАТУРЫ
- Khanferyan RA, Vybornaya KV, Radzhabkadiev RM, Evstratova VS, Nalivayko NV, Semin VB, et al. Frequency of consumption of sweet carbonated drinks by the population of different age groups of the Russian Federation. Problems of Nutrition. 2017;86(3):55–58. (In Russ.).
- Petrov AN, Galstyan AG, Radaeva IA, Turovskaya SN, Illarionov EE, Semipyatniy VK, et al. Indicators of quality of canned milk: Russian and international priorities. Foods and Raw Materials. 2017;5(2):151–161. DOI: https://doi.org/10.21603/2308-4057-2017-2-151-161.
- Bessonov VV, Khanferyan RA, Galstyan AG, Kucherov YuN. Potential side effects of caffeine consumption in healthy adults, pregnant women, adolescents and children (review of foreign literature). Problems of Nutrition. 2017;86(6):21–28. (In Russ.).
- Blaauboer BJ, Boobis AR, Bradford B, Cockburn A, Constable A, Daneshian M, et al. Considering new methodologies in strategies for safety assessment of foods and food ingredients. Food and Chemical Toxicology. 2016;91:19–35. DOI: https://doi.org/10.1016/j.fct.2016.02.019.
- Yu R, Yang B, Cai L, Lu X, Wang X. Excess free fructose beverages and allergy in children and adolescents: results from NHANES 2005–2006. Annals of Family Medicine. 2018;16(5):408–418. DOI: https://doi.org/10.1370/afm.2292.
- Hung H-C, Joshipura KJ, Jiang R, Hu FB, Hunter D, Smith-Warner SA, et al. Fruit and vegetables intakeand risk of major chronic disease. Journal of the National Cancer Institute. 2004;96(21):1577–1584. DOI: https://doi.org/10.1093/jnci/djh296.
- Farvid MS, Chen WY, Michels KB, Cho E, Willett WC, Eliassen AH. Fruit and vegetable consumption in adolescence and early adulthood and risk of breast cancer: population based cohort study. BMJ. 2016;353.
- Khan N, Afaq F, Kweon M-H, Kim K, Mukhtar H. Oral consumption of pomegranate fruit extract inhibits growth and progression of primary lung tumors in mice. Cancer Research. 2007;67(7):3475–3482. DOI: https://doi.org/10.1158/0008-5472.CAN-06-3941.
- Stowe CB. The effects of pomegranate juice consumption on blood pressure and cardiovascular health. Complementary Therapies in Clinical Practice. 2011;17(2):113–115 DOI: https://doi.org/10.1016/j.ctcp.2010.09.004.
- Singh M, Arseneault M, Sanderson T, Murthy V, Ramassamy C. Challenges for research on polyphenols from foods in Alzheimer’s disease: bioavailability, metabolism, and cellular and molecular mechanisms. Journal of Agricultural and Food Chemistry.2008;56(13):4855–4873.
- Gil MI, Tomás-Barberán FA, Hess-Pierce B, Holcroft DM, Kader AA. Antioxidant activity and total phenolic, organic acid and sugar content in commercial pomegranate juice. Journal of Agricultural and Food Chemistry. 2000;48(10):4581–4589.
- Pepin A, Stanhope KL, Imbeault P. Are fruit juices healthier than sugar-sweetened beverages? A review. Nutrients. 2019;11(5). DOI: https://doi.org/10.3390/nu11051006.
- Durnev AD, Oreshchenko AV. Pishcha: mutagenez i antimutagenez [Food: mutagenesis and antimutagenesis]. Storage and Processing of Farm Products. 1996;(3):14–18. (In Russ.).
- Büchner FL, Bueno-de-Mesquita HB, Linseisen J, Boshuizen HC, Kiemeney LALM, Ros MM, et al. Fruits and vegetables consumption and the risk of histological subtypes of lung cancer in the European prospective investigation into cancer and nutrition (EPIC). Cancer Causes and Control. 2010;21(3):357–371. DOI: https://doi.org/10.1007/s10552-009-9468-y.
- Rietjens IMCM, Martena MJ, Boersma MG, Spiegelenberg W, Alink GM. Molecular mechanisms of toxicity of important food-borne phytotoxins. Molecular Nutrition and Food Research. 2005;49(2):131–158. DOI: https://doi.org/10.1002/mnfr.200400078.
- DeChristopher LR, Uribarri J, Tucker KL. Intakes of apple juice, fruit drinks and soda are associated with prevalent asthma in US children aged 2–9 years. Public Health Nutrition. 2016;19(1):123–130. DOI: https://doi.org/10.1017/S1368980015000865.
- Hallfrisch J, Ellwood K, Michaelis OE, Reiser S, Prather ES. Plasma fructose, uric acid, and inorganic phosphorus responses of hyperinsulinemic men fed fructose. Journal of the American College Nutrition. 1986;5(1):61–68.
- Reiser S, Smith Jr JC, Mertz W, Holbrook JT, Scholfield DJ, Powell AS, et al. Indices of copper status in humans consuming a typical American diet containing either fructose or starch. The American Journal of Clinical Nutrition. 1985;42(2):242–251. DOI: https://doi.org/10.1093/ajcn/42.2.242.
- Bray GA, Nielsen SJ, Popkin BM. Consumption of high-fructose corn syrup in beverages may play a role in the epidemic of obesity. The American Journal of Clinical Nutrition. 2004;79(4):537–543.
- DeChristopher LR, Uribarri J, Tucker KL. Intake of high-fructose corn syrup sweetened soft drinks, fruit drinks and apple juice is associated with prevalent arthritis in US adults, aged 20–30 years. Nutrition and Diabetes. 2016;6(3). DOI: https://doi.org/10.1038/nutd.2016.7
- Dragsted LO, Daneshvar B, Vogel U, Autrup HN, Wallin H, Risom L, et al. A sucrose-rich diet induces mutations in the rat colon. Cancer Research. 2002;62(15):4339–4345.
- Tappy L, Mittendorfer B. Fructose toxicity: is the science ready for public health actions? Current Opinion in Clinical Nutrition and Metabolic Care. 2012;15(4):357–361. DOI: https://doi.org/10.1097/MCO.0b013e328354727e.
- Samoilov AV, Suraeva NM, Zaitseva MV, Kurbanova MN, Stolbova VV. Comparative assessment of artificial sweeteners toxicity via express biotest. Health Risk Analysis. 2019;(2):83–90. (In Russ.).
- Shahmohammadi M, Javadi M, Nassiri-Asl M. An overview on the effects of sodium benzoate as a preservative in food products. Biotechnology and Health Sciences. 2016;3(3).
- Golka K, Kopps S, Myslak ZW. Carcinogenicity of azo colorants: influence of solubility and bioavailability. Toxicology Letters. 2004;151(1):203–210. DOI: https://doi.org/10.1016/j.toxlet.2003.11.016.
- Amin KA, Abdel Hameid H, Abd Elsttar AH. Effect of food azo dyes tartrazine and carmoisine on biochemical parameters related to renal, hepatic function and oxidative stress biomarkers in young male rats. Food and Chemical Toxicology. 2010;48(10):2994–2999. DOI: https://doi.org/10.1016/j.fct.2010.07.039.
- Eberechukwu SE, Amadikwa AA, Okechukwu MO. Effect of oral intake of sodium benzoate on some haematological parameters of wistar albino rats. Scientific Research and Essays. 2007;2(1):6–9.
- Jacob SE, Hill H, Lucero H, Nedorost S. Benzoate allergy in children – From foods to personal hygiene products. Pediatric Dermatology. 2016;33(2):213–215. DOI: https://doi.org/10.1111/pde.12759.
- Kitano K, Fukukawa T, Ohtsuji Y, Masuda T, Yamaguchi H. Mutagenicity and DNA-damaging activity caused by decomposed products of potassium sorbate reacting with ascorbic acid in the presence of Fe salt. Food and Chemical Toxicology. 2002;40(11):1589–1594. DOI: https://doi.org/10.1016/s0278-6915(02)00119-9.
- Moura AG, Santana GM, Ferreira PMP, Sousa JMC, Peron AP. Cytotoxicity of cheese and cheddar cheese food flavorings on Allim cepa L. root meristems. Brazilian Journal of Biology. 2016;76(2):439–443. DOI: https://doi.org/10.1590/1519-6984.20514.
- Pandey H, Kumar V, Roy BK. Assessment of genotoxicity of some common food preservatives using Allium cepa L. as a test plant. Toxicology Reports. 2014;1:300–308. DOI: https://doi.org/10.1016/j.toxrep.2014.06.002.
- Almeida PM, Araújo SS, Santos IRMR, Marin-Morales MA, Benko-Iseppon AM, Santos AV, et al. Genotoxic potential of leaf extracts of Jatropha gossypiifolia L. Genetics and Molecular Research. 2016;15(1). DOI: https://doi.org/10.4238/gmr.15017415.
- Kurbanova MN, Suraeva NM, Rachkova VP, Samoylov AV. Comparative study of indicators of toxic activity in the Allium-test. Agrarian Bulletin of the Urals. 2018;171(4):25–30. (In Russ.).
- Zhang H, Jiang Y, He Z, Ma M. Cadmium accumulation and oxidative burst in garlic (Allium sativum). Journal of Plant Physiology. 2005;162(9):977–984. DOI: https://doi.org/10.1016/j.jplph.2004.10.001.
- Kuluev BR. Regulators of cell division and proliferation in plants. Biomics. 2017;9(2):119–135. (In Russ.).
- Lastdrager J, Hanson J, Smeekens S. Sugar signals and the control of plant growth and development. Journal of Experimental Botany. 2014;65(3):799–807. DOI: https://doi.org/10.1093/jxb/ert474.
- Ma QX, Cao XC, Xie YA, Xiao H, Tan XL, Wu LH. Effects of glucose on the uptake and metabolism of glycine in pakchoi (Brassica chinensis L.) exposed to various nitrogen sources. BMC Plant Biology. 2017;17. DOI: https://doi.org/10.1186/s12870-017-1006-6.
- Ivanova NN, Khomich LМ, Perova IB. Apple juice nutritional profile. Problems of Nutrition. 2017;86(4):125–136. (In Russ.).
