

Аннотация
Introduction. The existing diversity of plant raw materials and products predetermine the prospects of studying their potential as sources of pectin substances. However all current classifications are either fragmented or inconsistent.Study objects and methods. Our theoretical ivestigation aimed to develop an adequate classification for all taxa of plant origin, as well as their tissues and derivatives as pectin-containing materials. We developed criteria for assessing transformation potential of the protopectin complex based on the mass fractions of biologically active non-uronide components, native water-soluble pectin, the protopectin complex, and pectin substances. Individual boundary conditions were based on individual pectin potential, protopectin fragmentation potential, and pectin isolation potential.
Results and discussion. Based on the boundary conditions, we defined an universal criterion space that included a set of points M in the coordinates expressed by three main criteria. According to individual boundary conditions, the criterion space was divided, or zoned, into four domains corresponding to protopectin fragmentation potential. They were characterized by: 1) lack of pectin potential, 2) ineffective protopectin fragmentation, 3) ineffective isolation of fragmentation products, and 4) effective isolation. Finally, we developed a generalized algorithm to determine the location of points M[μ1, μ2 , μ3 ] in the zoned criterion space, characterizing the plant tissue.
Conclusion. Our approach can be used to assess any plant tissue for its protopectin transformation potential, which determines the technological influence on its pectin potential. This approach is universal, i.e., applicable to both plant tissue and its derivatives.
Ключевые слова
Protopectin complex, potential, transformation, evaluation system, criterion spaceВВЕДЕНИЕ
Food technology is currently striving to maximize the potential of raw materials and use new, nontraditional sources of essential nutraceuticals and food components with biological (antioxidants, enterosorbents, etc.) and/or technological (thickeners, stabilizers, etc.) functional activity [1, 2]. The most promising way to achieve that is a biotechnological approach that makes use of both living cultures of microorganisms and isolated enzyme systems. When using isolated enzyme systems, this approach involves a multiple stage fragmentation of a native supramolecular complex of plant and/or animal cell walls into target components with a wide range of physicochemical and/or technological properties [3–5].
One of the methods within this approach is to activate the potential of a multicomponent polymer matrix of cell walls and intercellular spaces. This method has a limited use in processing agricultural raw materials. It mainly consists in partial or complete degradation (depolymerization) of its individual components to change the consistency or transparency of the final product, or to clear it of degradation products and improve its sensory characteristics. Most certainly, a targeted use of this polymer matrix is complicated by its highly heterogeneous components, a system of bonds between them, and highly entangled polymer chains [6]. Moreover, the heterogeneity of individual matrix components is a serious obstacle to controlling their properties during extraction [7, 8].
Pectin substances are among major carbohydrate biopolymers that have a wide variety of functional and technological characteristics [9, 10]. In a plant cell, they are represented by two main fractions – native water-soluble pectin and a native water-insoluble protopectin complex. The last one is the most valuable for transformation due to its molecular structure and composition [9].
The structure of cell walls in almost all terrestrial plants makes them a potentially good resource for the industrial production of pectin [6, 11, 12]. However, it is difficult to implement. Since the protopectin complex is a branched supramolecular structure incorporated into the cell wall, its transformation is mainly fragmentation into water-soluble polymers (soluble pectin). In addition, mass fractions of pectin substances and the protopectin complex may depend on the type, grade, and purpose of raw materials, their structure and phase of development, soil and weather conditions for their vegetation, as well as localization, duration and storage conditions, processing intensity, etc. [10, 13]. In this regard, the choice of a plant as a pectin-containing material should be determined by the purpose of its use.
Raw materials can be classified according to the size of their pectin potential – “high”, “medium,” and “small” (“low”, “insignificant”) [9, 10, 14]. The only fundamental approach to pectin production was offered by Donchenko in [15] and supplemented by Rodionova et al. in [19, 20] (works [16–18] are actualy based on [15]). Although this approach is rather fragmented, it can be used as a basis for developing a universal system that takes into account the native pectin potential of plant tissue.
The protopectin complex is a key object whose fragmentation enables us to use the biomass of a plant material as a source of pectin substances. Due to the presence of certain plant organisms, mainly a natively soluble fraction of pectin, biomass can be attributed to potential sources of pectin. On the other hand, the biomass of certain taxonomic elements may contain a small amount of pectin, which makes its use ineffective.
Therefore, we found it relevant to develop a clearcut classification of plant bio-resources into groups to determine the prospects of their use as pectin-containing raw materials.
In this regard, we aimed to develop a system of criteria for assessing the transformation potential of native complexes of plant carbohydrate biopolymers exemplified by pectin. To achieve this aim, we set the following objectives:
– working out criteria to assess the transformation potential of native plant biopolymers and the concept of their applicability, and
– developing a system of boundary conditions and an universal algorithm for classifying plant materials according to the transformation potential of their native pectin components.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
According to existing data, all plant materials can be classified into four main groups, namely:
– bio-resources with sufficient potential for protopectin
fragmentation and subsequent isolation of its products as
independent substances;
– bio-resources with sufficient potential for protopectin
fragmentation, but with insufficient potential for
isolation of its products;
– bio-resources with insufficient potential for
protopectin fragmentation, but with sufficient potential
for natively soluble pectin;
– bio-resources with no pectin potential.
On the one hand, this differentiation involves unifying plant characteristics and reducing them to certain generalized values. On the other hand, it involves dividing the domain of generalized values into four fixed zones. As we know, a universal tool for unifying an arbitrary set of source factors is a range of anonymized criteria reducible to a certain system with the use of boundary conditions [21, 22]. Thus, we can apply a criteria-based approach to fulfilling our objectives.
To be able to scale the criteria to determine clear boundary conditions, we used Harrington’s individual desirability function in its canonical form [23]:
![]()
where di is the dimensionless value of Harrington’s individual desirability function; bi0 is the constant; bi1 is the coefficient; and φi is the dimensionless operator of Harrington’s individual desirability function.
We introduced the first and second individual criteria for protopectin fragmentation potential among the main criteria to assess the native pectin potential.
Let us begin with the first criterion. According to [7, 8], the presence of pectin in the tissue or a certain amount of protopectin in the cell wall matrix is not sufficient for assessing the native pectin potential of plant tissue. The tissues of many plant organisms also contain a significant amount of organic and mineral components with valuable vitamins and antioxidant activity, pronounced aroma, micro- and macronutrient values, etc. [17]. They are also highly sensitive to active technological impact factors. During protopectin fragmentation, organic and mineral components can enter into uncontrolled interactions, resulting in a partial or complete loss of their biological potential. Therefore, when assessing the native pectin potential, we should take into account the presence of these biologically active components among other significant factors.
Thus, we decided a complex operator as an independent variable, taking into account mass fractions of protopectin and biologically active components in the tissue:
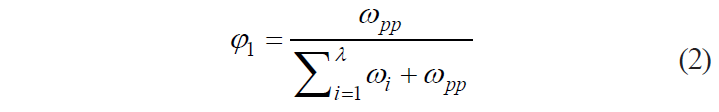
where ωpp is the mass fraction of protopectin, mg in 100 g; ωi is the mass fraction of the i-th biologically active component, mg/100 g; and λ is the number of biologically active components in the tissue (λ ∈ N).
To apply this operator in practice, we transformed it as follows:
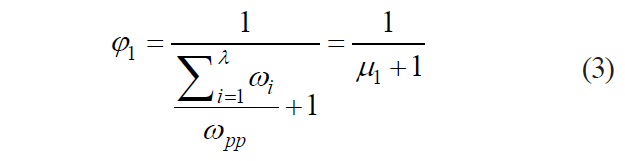
where
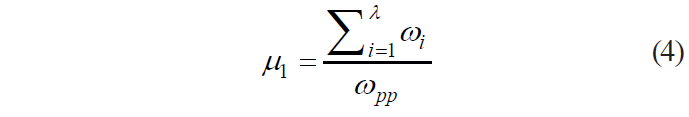
As we can see, with all possible values of ωpp and Σλi=1ωi, this criterion has the following range of definition:
![]()
In this case, Harrington’s individual desirability function can be expressed as:
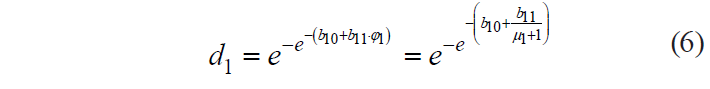
To determine the numerical values of b10 and b11, we had to set the primary relations between the pairs {μ11;d11} and {μ12 ;d12}, for which we proceeded from the following considerations.
If an i-th biologically active component has a specific measure of value pi, the total measure of value for all biologically active components under consideration is:
![]()
where vbac is the total measure of value for all biologically active components, units; mi is the mass of the i-th component, mg/100 g of plant tissue; m is the tissue mass, mg; pi is the specific measure of value of the i-th component, units/mg; and ωi is the mass fraction of the i-th component in the plant tissue, %.
If specific measures of value for the components are expressed through some average specific measure of value
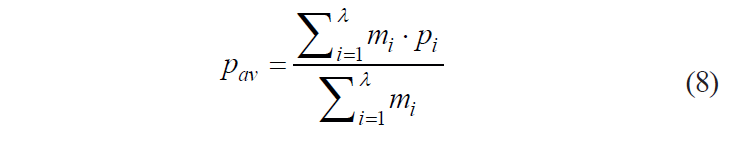
then formula (7) looks as follows:
![]()
from which
![]()
If we apply similar considerations to protopectin, then:
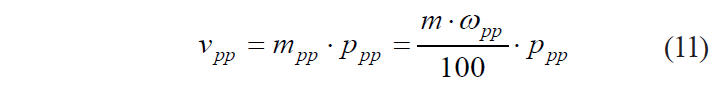
where vpp is the total measure of protopectin value, units; mpp if the mass of protopectin in the tissue, mg; ppp is the specific measure of protopectin value, units/mg; and ωpp is the mass fraction of the i-th component in the plant tissue, %.
From Eq. (11), it follows that
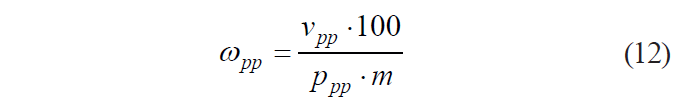
Thus, formula (4) can be presented as:
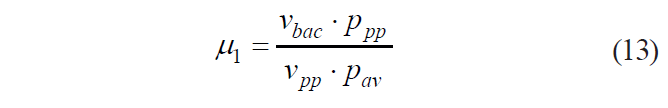
Grouping similar values on its sides, formula (13) can be transformed as:
![]()
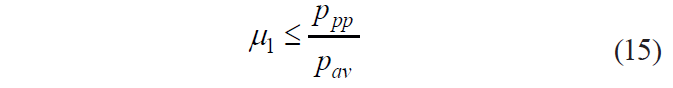
If pav is expressed as pi – in fractions of ppp, – then condition (15) looks as follows:
![]()
When calculating pav, it is advisable to use pi rather than pi, its value reduced to ppp:
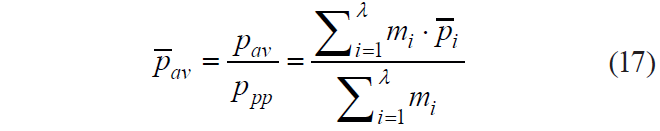
Theoretically, pi can be determined using several approaches. However, we believe that the most appropriate approach is based on a daily human need for individual nutrients. This approach is least opportunistic (compared to the financial approach) and subjective (compared to direct expert assessments). Naturally, daily requirements for certain components depend on our knowledge of biochemical processes in the human body, as well as on the constantly changing environmental situation in the world [24]. However, these factors should not significantly affect pav.
The value of pav was calculated in several stages.
At the first stage, we determined daily requirements for each of the biologically active components (ui) and pectin (ups) based on a daily energy requirement of 2000 kcal and an average body weight of 65 kg. The differences in daily requirements for men and women were averaged. For comparability, all the values were presented in mg/kg of body weight.
At the second stage, we calculated specific measures of value for biologically active components (pi) and pectin (pps):
![]()
![]()
The specific measures of value for pectin pps and protopectin ppp were numerically identical since protopectin is only valuable for the human body in the form of its fragmentation products. To simplify, we assumed that processing resulted in all protopectin fragmented in a targeted manner (i.e., into fragments that could be identified as pectin).
At the third stage, we determined specific measures of value in the fractions of the specific measure of pectin values pi.
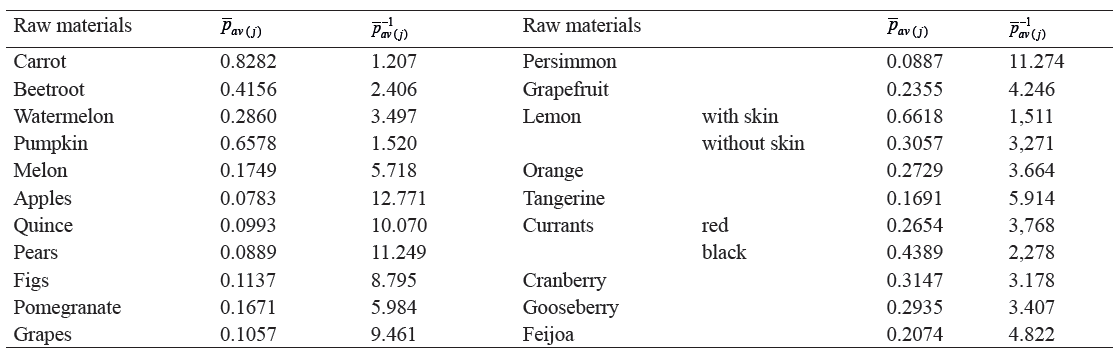
The calculation results are shown in Table 1.
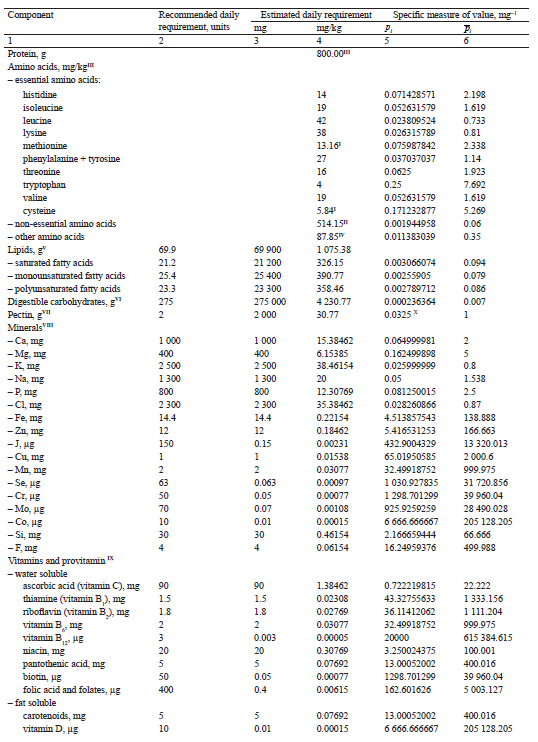
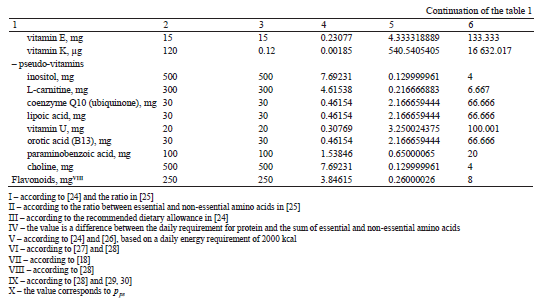
At the fourth stage, we calculated the value of p−1 av (Table 2). Based on the data in [31], we determined the content of biologically active components in 100 g of tissue for 21 types of plant materials from the classification presented in [16]. For each type of raw material, formula (17) was used to calculate the values of pav(j) and pav(j)−1, where j ∈ N.
Some assumptions were made in the calculations.
For example, the mass fractions of the components
which were not available in the database were assumed
as equal to zero [31]. The amount of carotenoids
was calculated based on the biological potential of
each type of raw material as ![]() , where mβ−car is the mass fraction of β-carotene, mg/100 g;
, where mβ−car is the mass fraction of β-carotene, mg/100 g; ![]() is the sum of mass fractions of
other carotenoids, mg/100 g [24]. The amount of
tocopherols was also calculated taking into account
the biological potential of each type of raw material
as
is the sum of mass fractions of
other carotenoids, mg/100 g [24]. The amount of
tocopherols was also calculated taking into account
the biological potential of each type of raw material
as ![]() , where mα−toc and mγ−toc are the
mass fractions of α- and γ-tocopherols, respectively;
mg/100 g [24]. To determine the sum of the remaining
amino acids, we subtracted the mass fractions of
essential and non-essential amino acids from the mass
fraction of protein.
, where mα−toc and mγ−toc are the
mass fractions of α- and γ-tocopherols, respectively;
mg/100 g [24]. To determine the sum of the remaining
amino acids, we subtracted the mass fractions of
essential and non-essential amino acids from the mass
fraction of protein.
The calculation results are shown in Table 2.
Since ![]() values were significantly different for different types of raw materials, we calculated the average
values were significantly different for different types of raw materials, we calculated the average ![]() and the margin of error Δ to determine boundary values (μ11 and μ12):
and the margin of error Δ to determine boundary values (μ11 and μ12):
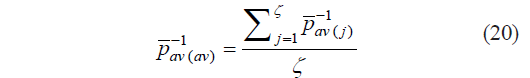
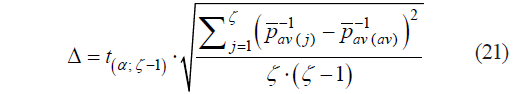
where ζ is the number of raw material types; t(α ;ζ−1) is Student’s t-test; and α is the probability of error (0.05).
Based on the above, the value of μ11 for the first pair {μ11;d11} was calculated as:
![]()
The value of μ12 for the second pair {μ12 ;d12} was calculated as the second order of μ11:
![]()
The critical (boundary) values of μ1 were based on the analysis of Harrington’s desirability function, using μ11 and μ12 as reference values. Since they are preset, the calculated values were rounded to the nearest whole number.
Despite the rigor of expression (16), its right-hand side is an empirical value based on the chemical composition of a finite number of plant raw materials and, therefore, it cannot be considered a priori. To make up for this feature, we further determined the critical values of μ1 on the basis of Harrington’s desirability function, using μ11 and μ12 as reference values.
Since a smaller reference value corresponded to a larger value of Harrington’s individual desirability function, we defined a condition Condd1 that determined the individual form of the function as:
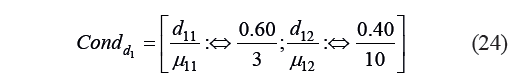
Based on Condd1, we calculated the values of the constant and the coefficient: b10 = −0.246; b11 = 3.673.
The critical values of the first criterion for the protopectin fragmentation potential at the points with standard critical values of the desirability function can be calculated using Eq. (6) with the variable μ1:
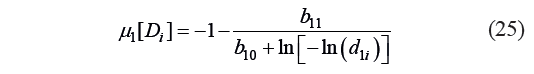
where μ1[Di] is the value of the criterion μ1 at the critical point Di of Harrington’s individual desirability function determined by Eq. (6) and corresponding to d1i; and d1i is the standard i-th critical (canonical) value d1 of Harrington’s individual desirability function.
The graphic interpretation of Harrington’s individual desirability function corresponding to the condition Condd1 is given in Fig. 1. For each value of d1i, we determined the corresponding values of μ1[Di].
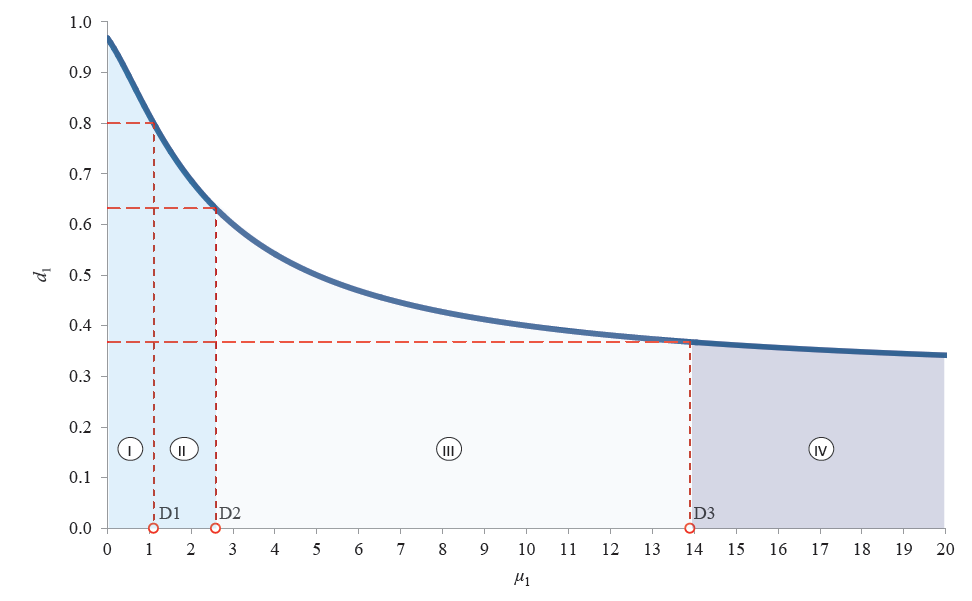
As we can see, the μ1 range of definition includes four domains separated by the critical values of μ1[Di], where i = 1, 2, 3. By definition, domain IV includes those μ1 values at which the fragmentation of the protopectin complex makes no sense due to a low value of the individual function of desirability.
Domain III covers those μ1 values at which the individual desirability function is large enough for protopectin fragmentation to make sense, but insufficiently large to neglect non-uronide bioactive components and isolate the products of fragmentation.
In domains I and II, the individual desirability function is so large that the content of non-uronide bioactive components in plant tissue can be completely ignored.
Based on the physical meaning of the boundary conditions for μ1, we established two individual boundary conditions that partially determined the native pectin potential of plant tissue.
Boundary condition I:
– μ1 > μ1[D3] means the absence of the first individual
potential for protopectin fragmentation;
– μ1 ≤ μ1[D3] means the presence of the first individual
potential for protopectin fragmentation.
Boundary condition II:
– μ1[D3] ≥ μ1 > μ1[D2] means the absence of the first
individual potential for isolation of protopectin
fragmentation products;
– μ1 ≤ μ1[D2] means the presence of the first individual
potential for isolation of protopectin fragmentation
products.
Next, we determined the structure and properties of the second dimensionless individual criterion for the protopectin fragmentation potential.
The second independent variable was a complex operator based on the mass fraction of protopectin in the tissue:
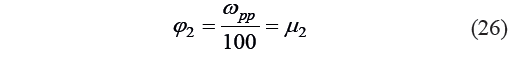
where φ2 is the dimensionless operator of Harrington’s individual desirability function; and μ2 is the second dimensionless individual criterion for the protopectin fragmentation potential.
Harrington’s individual desirability function was expressed as:
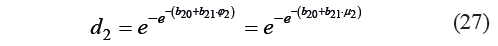
Thus, the condition Condd2 that determined the individual function was set as:
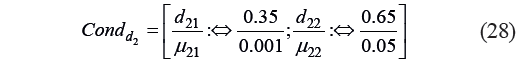
Based on Condd2, we calculated the values of the constant and the coefficient: b20 = −6.68×10−2 and b21 = 18.179. The critical values of the μ2 criterion were calculated as:
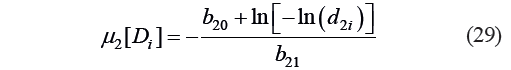
where μ2[Di] is the value of μ2 at the critical point Di of Harrington’s individual desirability function calculated by Eq. (6) and corresponding to d2i; d2i is the standard i-th critical (canonical) value d2 of Harrington’s individual desirability function.
The graphic interpretation of Harrington’s individual desirability function corresponding to the condition Condd2 is presented in Fig. 2. For each value of d2i, we calculated the corresponding values of μ2[Di].
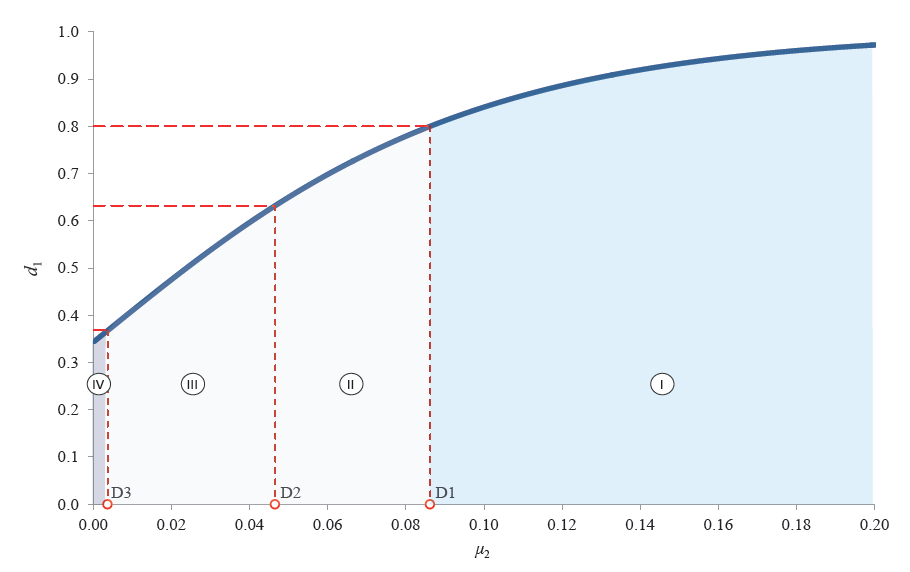
Just like with μ1, the μ2 range of definition includes four domains separated by the critical values of μ2[Di], where i = 1, 2, 3.
By definition, domain IV covers those values of μ2
at which the fragmentation of the protopectin complex
makes no sense. This led us to formulate the third
individual boundary condition:
– μ2 < μ2[D3] means the absence of the second
individual potential for protopectin fragmentation;
– μ2 ≥ μ2[D3] means the presence of the second individual potential for protopectin fragmentation.
We should note that fragmentation potentials I and II are categorical, i.e., if one of them is absent, the total fragmentation potential is absent as well.
Domains I, II, and III include such values of μ2 that
ensure not only protopectin fragmentation, but also
the isolation of fragmentation products. Based on the
canonical reference values of the individual desirability
function, we formulated the fourth boundary condition:
– μ2[D3] ≤ μ2 < μ2[D1] means the absence of the
second individual potential for isolation of protopectin
fragmentation products;
– μ2 ≥ μ2[D1] means the presence of the second individual potential for isolation of protopectin
fragmentation products.
Similar to the first and the second fragmentation potentials, the individual isolation potentials are categorical.
The third independent variable was a complex operator based on the mass fraction of pectin substances in the tissue:
![]()
where φ3 is the dimensionless operator of Harrington’s individual desirability function; ωps is the total amount of pectin substances, %; and μ3 is the third dimensionless individual criterion for the protopectin fragmentation potential.
In this case, the condition Condd3 that determined the individual function was calculated as:
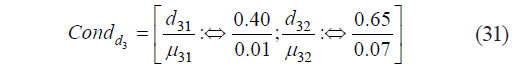
Based on expression (31), we calculated the constant and the coefficient as b30 = 3.8367×10−2 and b31 = 12.5788, respectively, and the critical boundaries of μ3, as:
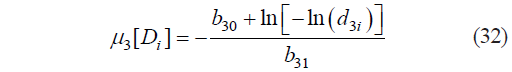
where μ3[Di] is the value of μ3 at the critical point Di of Harrington’s individual desirability function calculated by (6) and corresponding to d3i; and d3i is the standard i-th critical (canonical) value d3 of Harrington’s individual desirability function.
Figure 3 shows the graphic interpretation of Harrington’s individual desirability function given Condd3. For each value of d3i, we calculated the corresponding values of μ3[Di].

Here, we can clearly see domain IV with no pectin potential in the plant tissue.
As a result, we formulated the fifth individual
boundary condition:
– μ3 < μ3[D3] means the absence of pectin potential;
– μ3 ≥ μ3[D3] means the presence of pectin potential.
Thus, the pectin potential is categorical.
The fourth independent variable was a complex operator based on the ratio of the mass fractions of protopectin and pectin substances in the tissue:
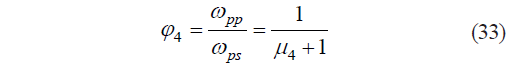
where φ4 is the dimensionless operator of Harrington’s individual desirability function; ωsp is the mass fraction of natively soluble pectin substances, %; and μ4 is the third dimensionless individual criterion for the protopectin fragmentation potential calculated as:
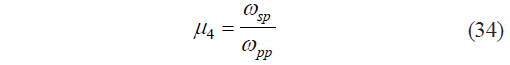
Then, the condition Condd4, which determined the individual function, was calculated as:
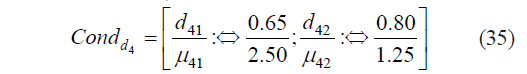
Based on expression (35), we calculated the constant and the coefficient (b40 = −0.3419, b41 = 4.1441).
Based on Condd4, the critical boundaries of μ4 were calculated as:
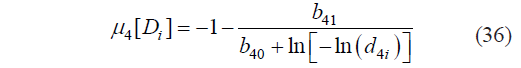
where μ4[Di] is the value of μ4 at the critical point Di of Harrington’s individual desirability function calculated by (6) and corresponding to d4i; and d4i is the standard i-th critical (canonical) value d4 of Harrington’s individual desirability function.
Figure 4 shows the graphic interpretation of Harrington’s individual desirability function given Condd4, with d4i values corresponding to μ4[Di] values.
Based on the logical content of d4i and the numerical values of μ4[Di], the range of definition can be divided into four domains that determine the fragmentation potential of the protopectin complex and the isolation potential of fragmentation products.
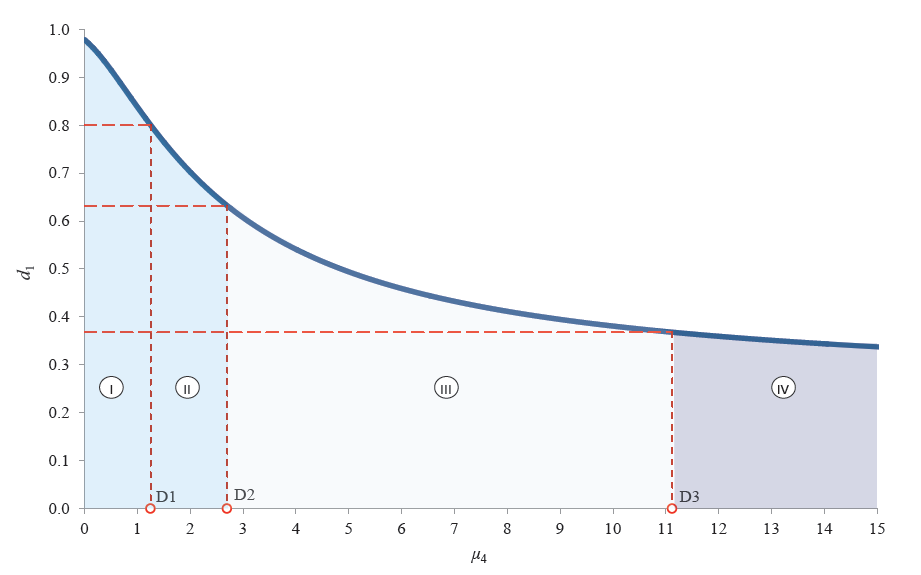
According to Fig. 4, domain IV covers those values
μ4 at which the mass fraction of water-soluble pectin
exceeds that of the protopectin complex so much
that there is practically no reason for its individual
fragmentation. Thus, we determined the sixth boundary
condition as follows:
– μ4 > μ4[D3] means the absence of the third individual
potential for protopectin fragmentation;
– μ4 ≤ μ4[D3] means the presence of the third
individual potential for protopectin fragmentation.
Following the same pattern, we determined the seventh
boundary condition (VII), namely:
– μ4[D2] < μ4 ≤ μ4[D3] means the absence of the
third individual potential for isolation of protopectin
fragmentation products;
– μ4 ≤ μ4[D1] means the presence of the third
individual potential for isolation of protopectin
fragmentation products.
In addition, boundary conditions VI and VII are based on:
![]()
where i = 3 for condition VI and, i = 2 for condition VII.
However, μ4 can be expressed as:
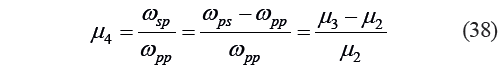
Then, given the presence of the third individual fragmentation potential:
![]()
Thus, the third individual potentials of fragmentation and isolation are relative since they are involved in the formation of respective total potentials indirectly, through expressions in which they act as one of the variables.
If we assume that there is a certain criterion space with coordinates μ1, μ2 and μ3, the pectin potential of any plant material can be clearly determined as a geometrical location of the point M[μ1, μ2, μ3] corresponding to the material under analysis.
Based on the a priori assumption that
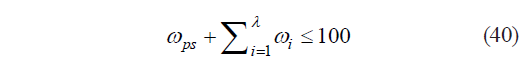
we can establish the eighth boundary condition (VIII): the top boundary of the range of definition for all possible values of M[μ1, μ2, μ3] is determined by the following basic proposition:
![]()
In addition, since a part cannot be larger than a whole, it is also true that:
![]()
which leads to the following condition:
![]()
i.e., the bottom boundary of the range of definition for all possible values of M[μ1, μ2, μ3] is determined by the second basic proposition:
![]()
The last formula is an expression of boundary condition IX.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
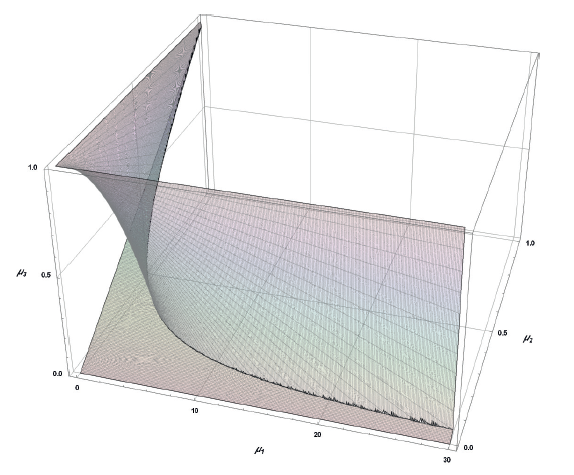
Thus, according to boundary conditions VIII and IX, a set (A) of all points M[μ1, μ2, μ3] can be defined as
![]()
graphically presented in Fig. 5.
The logic of assessing plant bioresources for the
presence of pectin substances determines general
boundary conditions for defining a set of points
M[μ1, μ2, μ3] as the following hierarchy: “individual pectin potential → individual fragmentation potential of
the protopectin complex → individual isolation potential
of protopectin fragmentation products”. Thus, the entire
set of points M[μ1, μ2, μ3] can be divided into four
subsets:
– subset A1 characterized by the absence of a common
pectin potential in all the elements;
– subset A2 where A2 ∩ A1 = ∅ and all the elements have
a common pectin potential, but lack a common potential
for protopectin fragmentation;
– subset A3 where A3 ∩ A2 = ∅ and all the elements
have common pectin and protopectin fragmentation
potentials, but lack a common isolation potential for
fragmentation products; and
– subset A4 where A4 ∩ A3 = ∅ and all elements
have common pectin and protopectin fragmentation
potentials, as well as isolation potential for
fragmentation products.
By definition, the following is true for all the subsets:
![]()
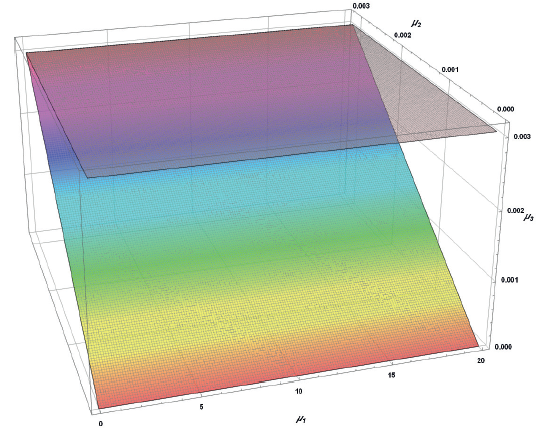
Based on the above, the existence of A1 corresponds to:
![]()
The area of definition for all A1 elements is partially presented in Fig. 6.
The existence of subset A2 corresponds to:
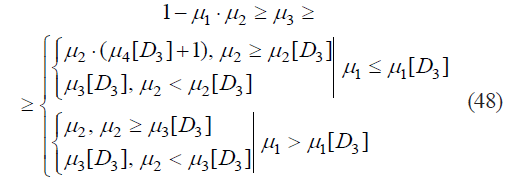
Figure 7 shows a partial area of definition for all A2 elements.
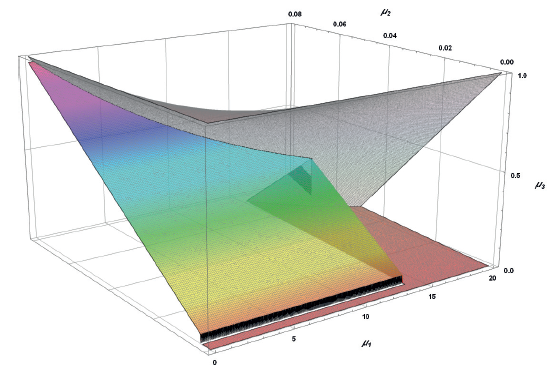
The existence of A3 corresponds to:

Figure 8 presents the area of definition for all A3 elements.
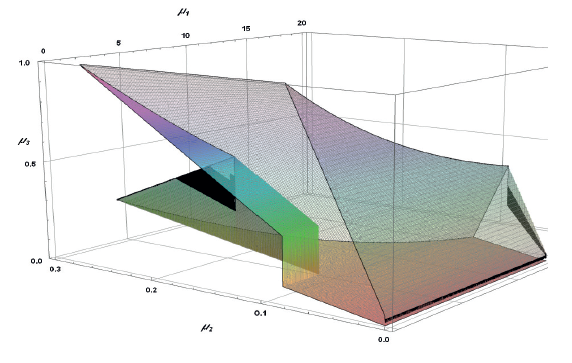
The existence of subset A4 corresponds to:
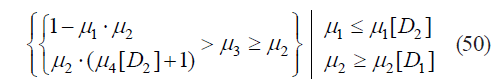
The area of definition for all A4 elements is presented in Fig. 9.
Thus, the specific value M[μ1, μ2, μ3] that shows its belonging to one of the subsets Ai (where i = 1, 2,3, 4) in the zoned criterion space clearly determines the plant tissue’s overall potential for protopectin fragmentation.
Our approach to classifying plants as pectincontaining materials, which is based on a system of criteria and a zoned criterion space, has clear advantages over existing methods due to its objectivity determined by the boundary conditions.
However, when analyzing this approach, we can
easily see that the μj1 and μj2 values corresponding
to dj1 and dj2 in the conditions ![]() were set
a priori, based on general assumptions regarding the
degree of acceptability of certain μj values within
Harrington’s individual desirability functions in
accordance with the boundary (canonical) values of d.
Yet, the conditions
were set
a priori, based on general assumptions regarding the
degree of acceptability of certain μj values within
Harrington’s individual desirability functions in
accordance with the boundary (canonical) values of d.
Yet, the conditions ![]() determine the coefficients and constants, and, consequently, individual desirability functions, as well a s n umerical values of μj[Di].
Therefore, at this stage, our approach has a general,
conceptual form requiring further research.
determine the coefficients and constants, and, consequently, individual desirability functions, as well a s n umerical values of μj[Di].
Therefore, at this stage, our approach has a general,
conceptual form requiring further research.
Based on the results, we developed a generalized algorithm to determine the geometric location of plant tissue in the zoned criterion space, or M[μ1, μ2, μ3] belonging to one of the subsets (Fig. 10). We can use this algorithm to assess any plant tissue’s potential for transformation of the protopectin complex, which determines the influence of any technological impact on its pectin potential.
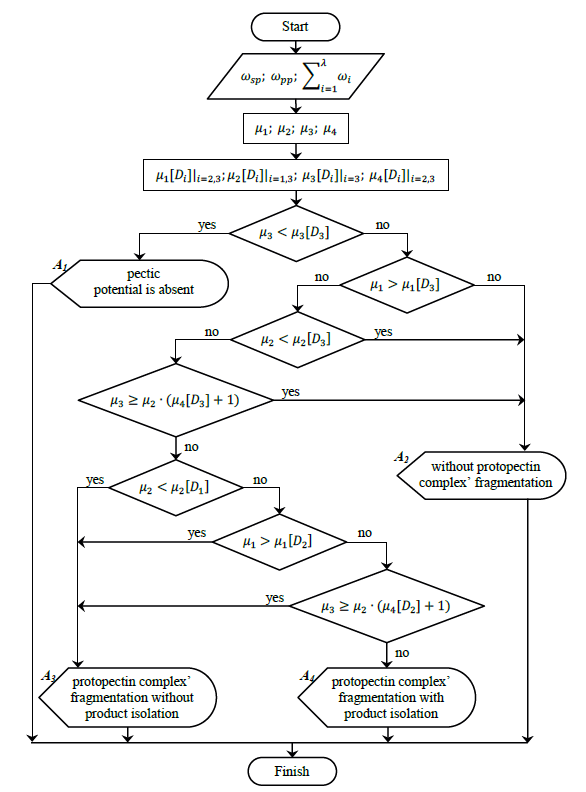
The approach that we used to determine the criterion space and boundary conditions for its zoning explicitly suggests that this algorithm is universal for classifying plant tissue or its derivatives as pectin-containing materials. Thus, the algorithm is applicable to any type of plant material for which the μ1, μ2 and μ3 criteria can be numerically expressed.
ВЫВОДЫ
To sum up, our investigation showed the following results.
1. We developed a system of criteria to assess the transformation potential of the protopectin complex in plant tissue. This system is based on the geometrical location of M[μ1, μ2, μ3] – the p oint that corresponds to the material under analysis – in a zoned criterion space with coordinates in the form of dimensionless individual criteria for protopectin fragmentation potential.
2. The dimensionless individual criteria for protopectin fragmentation potential included the ratio between the mass fractions of biologically active components and protopectin in plant tissue, the mass fraction of the protopectin complex expressed in unit fractions, and the mass fraction of total pectin substances expressed in unit fractions.
3. We established nine individual boundary conditions, individual pectin potential, two individual fragmentation potentials, and three individual isolation potentials for pectin substances, which altogether determine a system of zoning the criterion space.
4. The boundary conditions in the definition area for a set of points M[μ1, μ2, μ3] had the following hierarchy: individual pectin potential → individual fragmentation potential of the protopectin complex → individual isolation potential of protopectin fragmentation products.
5. We developed an algorithm to classify plant tissues according to protopectin fragmentation potential based on the geometric location in the zoned criterion space.
КОНФЛИКТ ИНТЕРЕСОВ
The authors state that there is no conflict of interest.ФИНАНСИРОВАНИЕ
The materials were prepared as part of the government assignment to Gorbatov Federal Scientific Center for Food Systems at Russian Academy of Sciences.СПИСОК ЛИТЕРАТУРЫ
- Galstyan AG, Aksyonova LM, Lisitsyn AB, Oganesyants LA, Petrov AN. Modern approaches to storage and effective processing of agricultural products for obtaining high quality food products. Herald of the Russian Academy of Sciences. 2019;89(2):211–213. DOI: https://doi.org/10.1134/S1019331619020059.
- Galstyan AG, Turovskaya SN, Ryabova AE, Illarionova EE, Semipyatnyi VK, Radaeva IA, et al. Technological additives as an element of dry milk properties directed formation. News of the National Academy of Sciences of the Republic of Kazakhstan. Series of Geology and Technical Sciences. 2019;4(436):95–102. DOI: https://doi.org/10.32014/2019.2518-170X.102.
- Lee BH. Fundamentals of food biotechnology. Wiley-Blackwell; 2015. 544 p. DOI: https://doi.org/10.1002/9781118384947.
- Bhatia SC. Food biotechnology. CRC Press; 2017. 412 p.
- Holban AM, Grumezescu AM. Preface for volume 14: Advances in biotechnology for food industry. In: Holban AM, Grumezescu AM, editors. Advances in biotechnology for food industry. Elsevier; 2018. pp. 23–26. DOI: https://doi.org/10.1016/B978-0-12-811443-8.00022-0.
- Caffall KH, Mohnen D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydrate Research. 2009;344(14):1879–1900. DOI: https://doi.org/10.1016/j.carres.2009.05.021.
- Thakur BR, Singh RK, Handa AK, Chemistry and uses of pectin – A review. Critical Reviews in Food Science and Nutrition. 1997;37(1):47–73.
- Srivastava P, Malviya R. Sources of pectin, extraction and its applications in pharmaceutical industry – an overview. Indian Journal of Natural Products and Resources. 2011;2(1):10–18.
- May CD. Industrial pectins: Sources, production and applications. Carbohydrate Polymers. 1990;12(1):79–99. DOI: https://doi.org/10.1016/0144-8617(90)90105-2.
- Müller-Maatsch J, Bencivenni M, Caligiani A, Tedeschi T, Bruggeman G, Bosch M, et al. Pectin content and composition from different food waste streams in memory of Anna Surribas, scientist and friend. Food Chemistry. 2016;201:37–45. DOI: https://doi.org/10.1016/j.foodchem.2016.01.012.
- Ovodov YuS. Current views on pectin substances. Russian Journal of Bioorganic Chemistry. 2009;35(3):269–284. DOI: https://doi.org/10.1134/S1068162009030017.
- Pectin [Internet]. [cited 2020 Mar 04]. Available from: https://en.wikipedia.org/w/index.php?title=Pectin&oldid=940586485.
- Sato MF, Rigoni DC, Canteri MHG, Petkowicz CLO, Nogueira A, Wosiacki G. Chemical and instrumental characterization of pectin from dried pomace of eleven apple cultivars. Acta Scientiarum – Agronomy. 2011;33(3):383–389. DOI: https://doi.org/10.4025/actasciagron.v33i3.7125.
- Baker RA. Reassessment of some fruit and vegetable pectin levels. Journal of Food Science. 1997;62(2):225–229. DOI: https://doi.org/10.1111/j.1365-2621.1997.tb03973.x.
- Donchenko LV. Razrabotka i intensifikatsiya tekhnologicheskikh protsessov polucheniya pektina iz sveklovichnogo i drugikh vidov syrʹya [Development and intensification of technological processes for the production of pectin from beet and other raw materials]. Dr. eng. sci. diss. Kiev, 1990. 360 p.
- Donchenko LV, Karpovich NS, Simkhovich EG. Proizvodstvo pektina [Pectin production]. Kishinev: Shtiintsa; 1994. 181 p. (In Russ.).
- Donchenko LV. Tekhnologiya pektina i pektinoproduktov [Technology of pectin and pectin products]. Moscow: DeLi; 2000. 256 p. (In Russ.).
- Donchenko LV, Firsov GG. Pektin: osnovnye svoystva, proizvodstvo i primenenie [Pectin: basic properties, production and application]. Moscow: DeLi print; 2007. 275 p. (In Russ.).
- Rodionova LYa. Teoreticheskoe i ehksperimentalʹnoe obosnovanie tekhnologii pektinosoderzhashchikh izdeliy funktsionalʹnogo naznacheniya [Theoretical and experimental substantiation of the technology of pectin-containing functional products]. Dr. eng. sci. diss. Krasnodar: Kuban State Technological University; 2004. 48 p.
- Rodionova LYa, Donchenko LV, Sobol IV, Stepovoy AB. Pectin containing raw materials classification extension. Proceedings of the Kuban State Agrarian University. 2015;(52):199–206. (In Russ.).
- Galstyan AG, Semipyatnyy VK. K voprosu o rasshirenii oblasti otsenochnykh kriteriev kachestva pishchevykh produktov [On the issue of expanding the field of evaluation criteria for food quality]. Aktualʹnye voprosy industrii napitkov [Current issues in the beverage industry]. 2017;(1):27–29. (In Russ.).
- Oganesyants LA, Khurshudyan SA, Galstyan AG, Semipyatny VK, Ryabova AE, Vafin RR, et al. Base matrices – Invariant digital identifiers of food products. News of the National Academy of Sciences of the Republic of Kazakhstan. Series of Geology and Technical Sciences. 2018;6(432):6–15. DOI: https://doi.org/10.32014/2018.2518-170X.30.
- Harrington EC. The desirability function. Industrial Quality Control. 1965;21(10):494–498.
- Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein, and amino acids. Washington: National Academies Press; 2005. 1358 p. DOI: https://doi.org/10.17226/10490.
- Protein and amino acid requirements in human nutrition: report of a joint FAO/WHO/UNU expert consultation. Geneva: World Health Organization; 2007. 265 p.
- Fats and fatty acids in human nutrition: report of an expert consultation. Rome: FAO; 2010. 166 p.
- Carbohydrates in human nutrition. Report of a Joint FAO/WHO Expert Consultation. Rome: FAO; 1998. 140 p.
- MR 2.3.1.2432–08 Normy fiziologicheskikh potrebnostey v ehnergii i pishchevykh veshchestvakh dlya razlichnykh grupp naseleniya Rossiyskoy Federatsii [Norms of physiological requirements for energy and nutrients for various population groups of the Russian Federation]. Moscow: Federal Center for Hygiene and Epidemiology of Rospotrebnadzor; 2009. 36 p.
- Vitamin and mineral requirements in human nutrition. 2nd ed. Rome: WHO and FAO; 2004. 341 p.
- Human vitamin and mineral requirements. Rome: FAO; 2001. 303 p.
- U.S. Department of Agriculture [Internet]. [cited 2020 Mar 04]. Available from: https://fdc.nal.usda.gov.
