

Аннотация
Introduction. Due to the trend of avoiding antibiotics and acquiring eco-friendly products, the use of environmentally safe preparations is becoming increasingly relevant in poultry farming.Study objects and methods. We used Salmonella enteritidis and Campylobacter jejuni isolated from poultry carcasses. At the first in vitro stage, we studied the ability of mannan oligosaccharides, isolated from the cell walls of Saccharomyces cerevisiae yeast, to adsorb bacterial pathogens. At the second stage, we studied the influence of fraction on the activity, colonization and microflora composition of ducklings’ intestines. At the third stage, we determined the antagonistic activity of Bifidobacterium spp. (Bifidobacterium lactis, Bifidobacterium longum, Bifidobacterium bifidum) and Lactobacillus spp. (Lactobacillus fermentun, Lactobacillus salivarius, Lactobacillus acidophilus) against Salmonella enteritidis and Campylobacter jejuni isolates. The experiment was conducted on the ducklings of Star 53 H.Y. cross. Their diet was supplemented with probiotics, prebiotics, and their combination.
Results and discussion. In vitro studies showed the ability of mannan oligosaccharides isolated from the cell walls of Saccharomyces cerevisiae yeast to adsorb Salmonella enteritidis and Campylobacter jejun. In vivo experiment showed the ability of mannan oligosaccharides to prevent colonization of poultry intestines by bacterial pathogens with type I fimbriae.
Conclusion. The reisolation rate of ducks infected with Salmonella enteritidis was 53.6% lower, and those infected with Campylobacter jejuni, 66.2% lower than the control. Mannan oligosaccharides added to the diet did not affect the concentration of lactobacilli, enterococci, and anaerobic bacteria in the ducks’ intestines. A combined use of Bifidobacterium spp. and mannan oligosaccharides improved the preservation of poultry stock by 8.7%, which made it an effective way to prevent poultry salmonellosis.
Ключевые слова
Prebiotics, probiotics, mannan oligosaccharides, microorganisms, bacterial pathogens, Salmonella spp., Campylobacter spp., poultry, ducks, productivityВВЕДЕНИЕ
In the world production of poultry, the share of waterfowl meat is 7.2%, specifically duck meat – 4.2%, goose meat – 3%. Their share in the gross production of poultry meat tends to increase. In industrial poultry farming, the problem of controlling bacterial infections of waterfowl is of genuine concern. Salmonella and Campylobacter are considered the most common etiological zoonotic factors worldwide, with productive poultry being the main source of infection.
In recent years, there has been an increase in the relative number of infections caused by Salmonella spp. and Campylobacter spp. The microorganisms are widespread in most warm-blooded and farm animals, including poultry. Ducks’ infection with salmonella can be detected at the age of about 14 days, and by the end of cultivation the whole flock can be found infected. Experimental studies showed that a small dose (less than 40 CFU) of S. enteritidis is sufficient to fully colonize the poultry intestines. This can lead to complete flock’s infection in 48 h [5–7]. Microorganisms can colonize the intestinal tract of poultry in large quantities, often at above 106–108 CFU/g of intestinal contents. The highest concentrations of bacterial pathogens are known to be present in the intestinal mucosa [4].
Poultry products can be contaminated at many stages of the “from farm to table” food chain, but the strategic one is the stage of primary poultry production. Following biosafety guidelines of GMP/HACCP significantly reduces the colonization of poultry by bacterial pathogens and, later, the contamination of carcasses during processing. The European Food Safety Agency’s monitoring (2008–2018) showed that about 86% of poultry carcasses in Europe were contaminated with Campylobacter and Salmonella bacteria.
In poultry production, main methods of infection control are taken at the stage of the cultivation in farms. Environmentally safe methods that ensure poultry quality and safety hold promise. Effective systems of poultry cultivation, feeding, and maintenance are reqired to control the spread of Salmonella and Campylobacter in poultry products. Bio-safety measures, decontamination of dropping and water are potentilly productive. Antibacterial drugs in treating of bacterial infections in poultry are considered a risk factor contributing to the development of antibioticresistant strains.
Following the trend of avoiding antibiotics, the search for new control methods is becoming increasingly important in poultry farming. The application of antimicrobial alternatives is highly potential. They include feed additives that are inhibitors of bacterial pathogens, as well as probiotics, prebiotics, bacteriophages, bacteriocins, which in combination prevent antibiotic-resistant strains of microorganisms and inhibit their proliferation [9–12].
Consequently, natural alternative antibacterial preparations are a way to reduce poultry gut colonization by pathogenic microflora. This is the most acceptable natural alternative to salmonella and campylobacter control that is economically viable and does not pose a risk to human health, animals, or the environment [3, 9]. Effective protection of poultry against pathogens, naturalness and safety, growth promotion, and economic effectiveness are the criteria for new alternatives to antibiotics [11, 13].
One of the requirements for probiotics use is the competitiveness of antagonistic microflora found in them. In order to prevent intestinal colonization by bacterial pathogens, probiotics are recommended for use from the first day of the birds’ life. Prebiotics promote the development of birds’ own symbiotic microflora, which can inhibit pathogens and reduce their adhesion to enterocytes.
Research suggests that some natural compounds have biological activity against salmonella proliferation, but few have shown efficacy in experiments on animals. “Actigen” prebiotic (Alltech) is a concentrated pure fraction of mannan oligosaccharides isolated from the cell walls of Saccharomyces cerevisiae yeast. The main advantage of these complex carbohydrates is their ability to adsorb certain strains of bacteria that have type I fimbriae (mannose-sensitive) and prevent intestinal colonization by pathogens. Besides, the industrial experiment proved the influence of combined use of mannan oligosaccharides and probiotics on intestinal microbiocenosis and duck productivity [14, 15].
We aimed to develop a method for preventing bacterial infections and increasing duck productivity using probiotics and prebiotics. The method was based on the study of adsorbing capacity of mannan oligosaccharides (MOS) and antagonistic properties of Bifidobacterium spp. and Lactobacillus spp. against Salmonella enteritidis and Campylobacter jejuni. We also aimed to analyze a combined effect of the cultures on gut microbiocenosis (activity and colonization) and on productivity of ducks.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
We used Salmonella enteritidis and Campylobacter jejuni isolated from poultry carcasses of Ukrainian farms. The studies were carried out in 2014–2018 at Sumy National Agrarian University, Sumy. The poultry carcasses were subjected to a detailed examination for pathomorphological changes. The liver, muscles, cloaca contents, ovaries, and various segments of the ovoid were aseptically assembled to be screened for salmonellosis and campylobacteriosis. Isolation and identification of microorganisms was carried out using tests recommended by “Bergey’s Manual” (1997) [35].
At the first stage (in vitro), we studied the ability of mannan oligosaccharides isolated from the cell walls of Saccharomyces cerevisiae yeast to adsorb bacterial pathogens. In our experiments we used 27 strains of Salmonella enteritidis and 13 strains of Campylobacter jejuni isolated from ducks’ chilled carcasses (liver, muscles, cloaca).
We used the daily agar culture of bacteria with 1% red blood cells of guinea pigs. Salmonella (1.5×109 CFU/mL) was used as an antigen. Erythrocytes were derived from the blood of a pre-selected donor (guinea pigs). Blood was placed in flasks containing sodium citrate and filtered through a cotton gauze filter to remove fibrin and small blood clots. Blood was centrifuged with sodium chloride isotonic solution four times (1500 rpm, 10 min). Then we introduced it into a 10% suspension of phosphate buffer solution (pH 7.0–7.2). The washed red blood cells were stabilized with 0.2% acrolein (acrylic aldehyde) solution in the phosphate buffer (1:1) and incubated in water bath at 37°C for 30–40 min while stirring periodically. Erythrocytes were washed three times by centrifuging with phosphate buffer at 5000 rpm. To improve the sorption properties of red blood cells, we treated them with tannin, combining equal parts 5% of frozen stabilized red blood cells and tannin solution (1:30 000). The mixture was left in the thermostat at 37°C for 40 min, then it was washed twice with phosphate buffer solution (pH 7.2–7.4) and then twice with sodium chloride isotonic solution (pH 7.2–7.4). To sensitize the antigen, we used a 1% red blood cell suspension. Suspensions were left for 24 h at 4°C to exclude spontaneous hemagglutination.
The degree of agglutination of the salmonellas isolated was determined by combining prepared suspended microorganisms and the aqueous solution of mannan oligosaccharides (0.2, 0.3 and 0.4 g/L) in a ratio of 1:1. E. coli O2 test culture was used as a positive control of the agglutination level of the pathogen. One-percent red blood cell suspension in phosphate buffer solution (pH 7.2–7.4) was used as a negative control [16–18].
At the second stage, we studied the influence of fraction (Aktigen, Alltech Inc.) on the activity, colonization and species composition of the microflora of young ducks’ intestines. Sixty male ducklings aged 30 days were used in the study. Each experiment involved one control and two experimental groups (50 heads in each). First experimental group was infected with Salmonella enteritidis, and the other group with Campylobacter jejuni (1×104 CFU/mL per os). Ducklings were kept in sterile boxes on the floor and fed by standards. They had free access to feed and water. In experimental groups, the birds received a prebiotic fraction of MOS (0.4 kg/t) together with the feed. Ten days after the infection we determined the concentration of salmonellas, campylobacil, lactobacil, bifidobacterium, and total concentration of anaerobic bacteria using dilution plate counting.
At the third stage, we determined the antagonistic activity of Bifidobacterium spp. (1.0×109 CFU/mL): Bifidobacterium lactis, Bifidobacterium longum, Bifidobacterium bifidum and Lactobacillus spp. (1.0×109 CFU/mL): Lactobacillus fermentun, Lactobacillus salivarius, Lactobacillus acidophilus against Salmonella enteritidis and Campylobacter jejuni isolates. Suspensions of bacterial probiotic cultures in a concentration of 1×109 m.c/cm3 were sown on Petri dishes and incubated for 24 h at 37°C. After that, suspensions with microorganisms (Salmonella enteritidis and Campylobacter jejuni) in a concentration of 1×109 m.c/cm3 were inoculated by streaking. The dishes with inoculation were incubated at 37°C for 24–72 h. We recorded the diameter of zones with no growth of test cultures. To control microbial growth, we used Preston-agar for Campylobacter, “Salmonella different agar” for Salmonella, as well as MPA and MPB for probiotics.
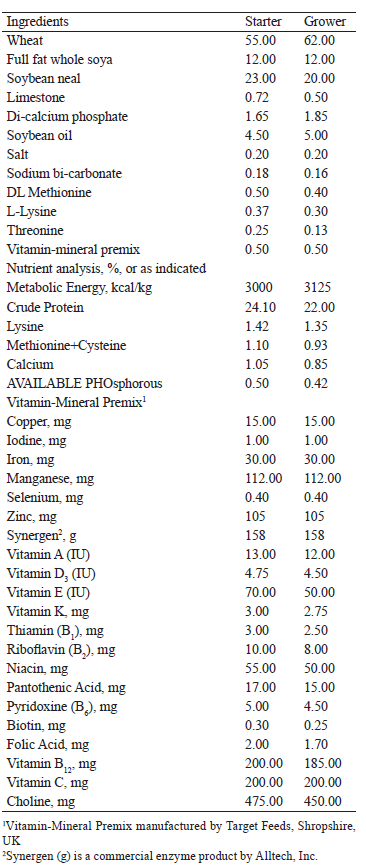
We used the Star 53 H.Y. cross ducklings to determine the effectiveness of probiotics, prebiotics, and their combination. The birds were randomly divided into 4 groups, 123 birds in each. Each group included 3 flocks, 41 birds in each (12 flocks in total). The control group received the main diet only. Three experimental groups received three different supplements in addition to the main diet: bifidobacteria (1.5×109 CFU/mL), mannan oligosaccharides (“Actigen” prebiotic), and a combination of Bifidobacterium spp. and Lactobacillus spp. (1.5×109 CFU/mL) in a ratio of 1:1 and the fractions of mannan oligosaccharides (0.4 kg/t of feed). These supplements were mannan-rich fractions isolated from the cell wall of Saccharomyces cerevisae yeast. The main diets were prepared at a commercial feed mill and consisted mainly of wheat and soybean flour, as shown in Table 1 [19, 20].
No principles of the bioethics code were violated during the experiments [21].
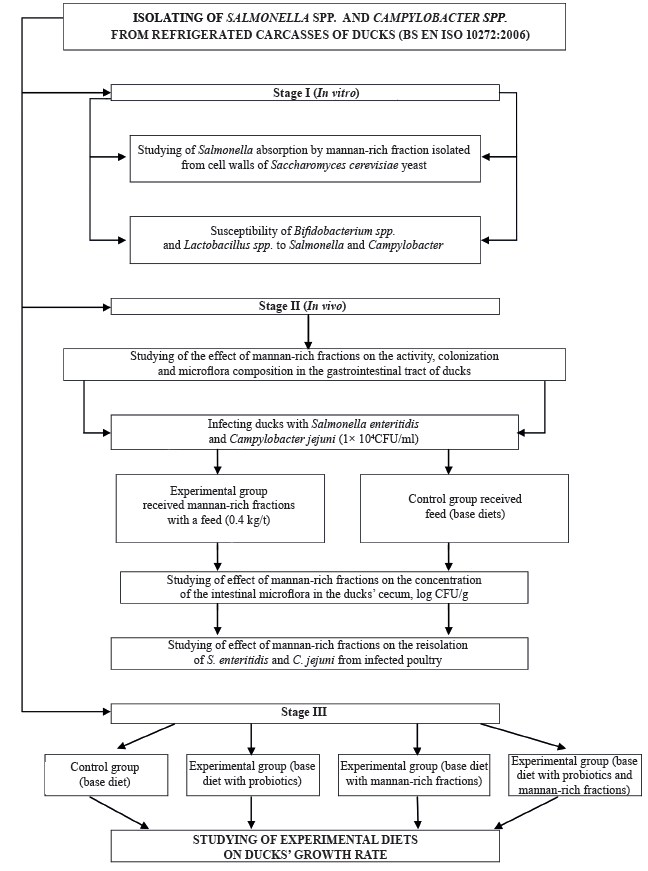
The general scheme of our experimental and practical studies is shown in Fig. 1.
Bacteriological analysis. We tested the laying houses for Campylobacter spp. before placing the birds there and on day 21. According to the methods described in BS EN ISO 10272:2006, the swabs were placed in 50 mL of isotonic solution and kept at 260 rpm for one minute [23, 24]. The suspension (0.1 cm3) was then transferred to two dishes with breeding ground (Preston agar base for Campylobacter) and incubated in microaerobic atmosphere (85% N2, 10% CO2 and 5% O2) at 40.5 ± 1°C. Then we examined them in 44 ± 4 h for typical and/or suspicious Campylobacter spp colonies.
Then, Salmonella enteritidis was isolated from the material. The serotyping of Salmonella spp. was carried out according to the methods with some modification according to the data [28, 32–34].
Statistical analysis. Weight gains and feed conversion rates were studied for statistical group differences using the Student’s T-test. The results of the microbiological analysis were logarithmic and evaluated for the statistical difference between the indicators that were measurable.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
The aim of our research was to study effects of mannan oligosaccharides fractions and probiotics on Salmonella enteritiadis and Campylobacter jejuni.
In vitro experiments showed that 0.2–0.4% aquatic fractions of mannan oligosaccharides could adsorb all the Salmonella strains and E. coli O2 test cultures (positive control).
We detected the most active and pronounced ability to adsorb bacterial pathogens in vitro experiments with 0.4% aqueous fraction of mannan oligosaccharides. We recorded the beginning of the adsorption process within 2 min. The active process was manifested in the form of finely-divided sediment and clearing of the supernatant. In 8–10 min we observed significant sedimentation (Fig. 2 a–d).

The formation of the sediment illustrates the adsorption process that occurred in the test tube. The same process can occur in the gut in animals and poultry.
Intestinal colonization by pathogens begins with the binding of cells to the epithelium of the intestinal mucosa [17]. Pathogens, including most types of Salmonella, E. coli, and Campylobacter attach to the gut via receptors (fimbriae) specific to certain carbohydrates containing mannose, which localize on the surface of intestinal mucosal epithelium cells [14].
When entering the intestines of poultry with feed, mannan-rich fractions bind to receptors of bacterial cells that have type I fimbriae (mannose-sensitive). Fractions of mannan oligosaccharides are not broken down by digestive enzymes and are held firmly on the surface of bacteria. Bacteria with blocked receptors cannot gain a foothold on the surface of epithelial cells – they transit through the gastrointestinal tract [13]. Thus, we found that the active concentration of mannan-rich fractions could successfully adsorb Salmonella, a pathogen that can cause foodborne diseases.
The following experiment examined the effects of fractions rich in mannanooligosaccharides on the activity, colonization, and species composition of microflora in ducks’ intestines.
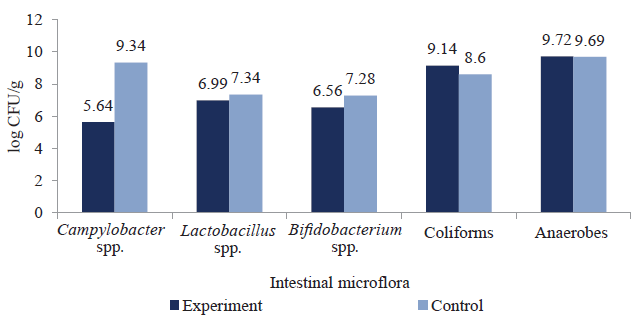
At the second (in vivo) stage, we determined the effect of mannan-rich fractions on the number of bacteria in the gut of experimentally infected ducks aged 30 days by type I fimbriae bacterial strains (C. jejuni and S. enteritidis strains). In experimental groups of birds that received prebiotic MOS fractions with feed, the level of bacteria with type I fimbriae decreased. The effect of mannan oligosaccharide-rich fractions on the concentration of intestinal microflora of ducks infected with S. enteritidis is shown in Fig. 3.
The effect of mannan-rich fractions on the concentration of intestinal microflora of ducks infected with C. jejuni is shown in Fig. 4.
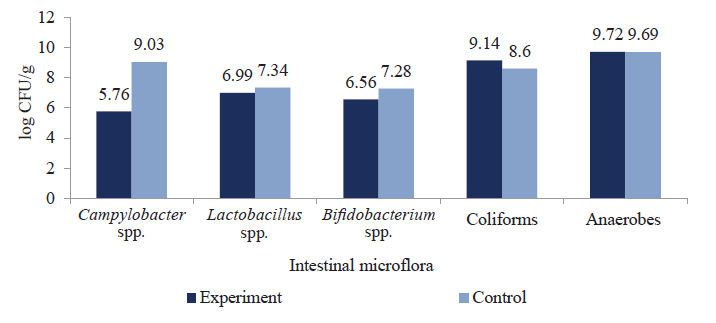
The results showed that mannan oligosaccharides could regulate intestinal microflora due to their selective ability to inhibit Salmonella spp. and Campylobacter spp. proliferation, preventing pathogenic colonization of the intestines and minimizing its toxic effect on the poultry. Concentration of Salmonella spp. in the ducklings’ gut was lower by 3.69 log CFU/g and Campylobacter spp. by 3.27 log CFU/g compared to the control, respectively. Metabolites of functional oligosaccharides did not affect the levels of intestinal colonization by pathogenic bacteria (coliforms and anoerobeds). They did not prevent Lactobacillus spp. and Bifidobacterium spp. proliferation either, which contributed to the colonization of beneficial bacteria in the birds’ intestines.
Regulation of intestinal microbiocenosis can potentially have a positive effect on immune response mechanisms, i.e. to strengthen immunity and enhance the poultry population.
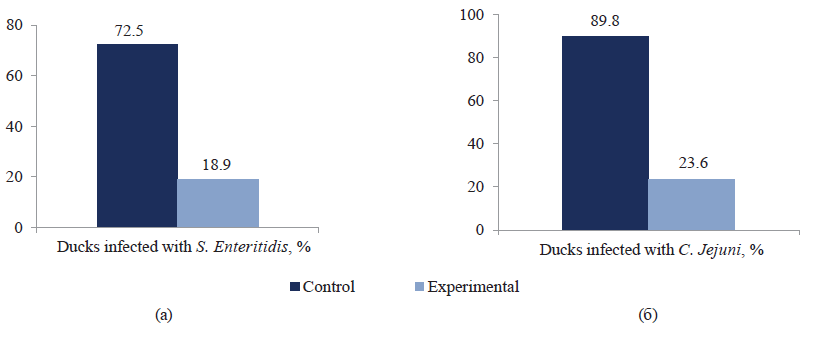
The effect of mannan-rich fractions on the ducklings’ gut microflora infected with S. enteritidis is shown in Fig. 4a. The reisolation rate of S. enteritidis and C. jejuni in the test group, which received prebiotic mannan-rich fractions with feed, decreased by 53.6 and 66.2%, respectively, compared to the control group (Figs. 5a, 5b).
Bifido- and lactobacteria also displayed antagonistic activity against Campylobacter jujuni and Salmonella enteritidis isolates. It makes them possible to be used for the prevention of infectious diseases caused by sensitive strains of pathogens to prebiotic drugs. Bifidobacterium spp. and Lactobacillus spp. suppressed the growth of microorganisms to different extents (Table 2).

Twelve isolates (92.6%) of Campylobacter spp. were susceptible to bifidobacteria. The inhibition zone of campylobacter was 5.1 ± 0.3 mm. Ten Campylobacter jujuni isolates showed a moderate level of antagonistic activity ‒ 76.9%, with the inhibition zone of 5.1 ± 1.0 mm.
Twenty four isolates (88.9%) of S. enteritidis were susceptible to bifidobacteria; the inhibition zone of S. enteritidis was 5.5 ± 0.4 mm. The antagonistic activity of lactobacilli against S. enteritidis showed a moderate level: 22 isolates (81.5%) had inhibition zone of 4.9 ± 0.5 mm. Bifidobacteria were more active against Campylobacter spp. and Salmonella spp. It makes it possible to use probiotics to prevent and treat infectious diseases caused by susceptible strains of pathogenic microorganisms to the drug. To improve the ducks’ productivity, we studied the effect of mannan-rich fractions. The experiment plan is given in Table 3.
To solve the problem of bacteriosis prevention and increase of birds’ productivity, we also studied the effect of a combined use of mannan oligosaccharides and probiotic bifidobacteria and lactobacilli.
The first stage of the experiment included 20320 ducks (Star 53 Y.Y.) divided into four groups: one control and three experimental ones. The experiment was carried out three times (81280 ducks in total). Probiotics were added to the diet of the ducks with water (10 cm3 per 4 kg duck weight) once a day from the first day until the end of the fattening period (49 days).
We added mannan-rich fractions to the base diet – 400 g/t of feed. We added bifidobacteria and mannanrich fractions to the duck diet once a day until the end of fattening period (49 days). The analysis showed higher results during all periods of the birds’ life compared to the control groups (Table 4).
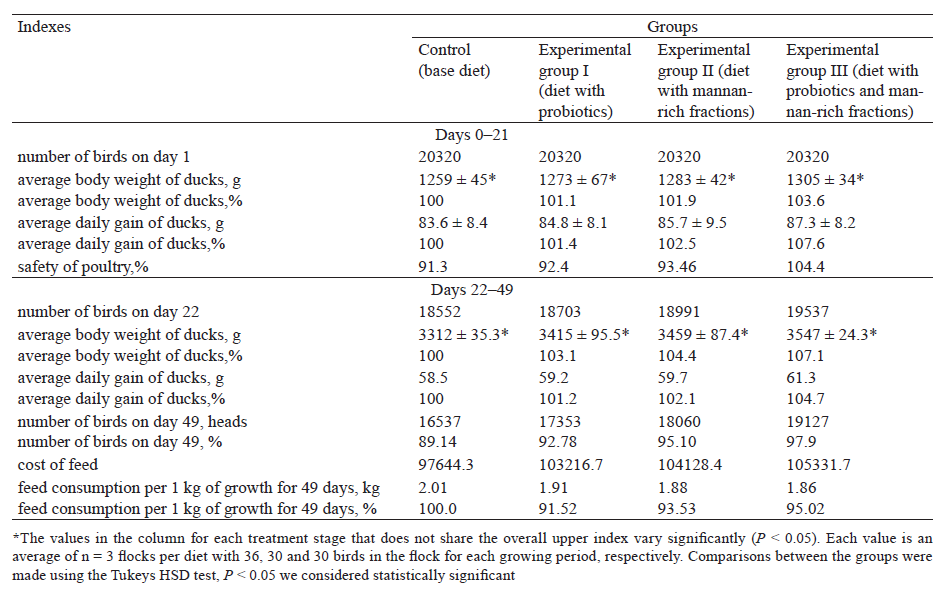
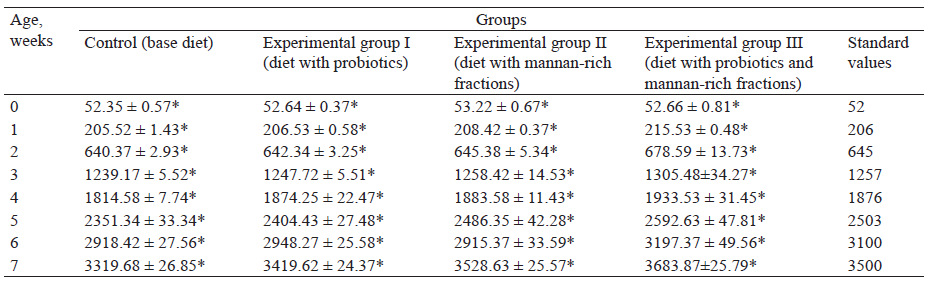
At the age of 21 days, the average growth rate of ducks receiving probiotics with mannan-rich fractions was 87.3 g vs. to 83.6 g in the control group. We noticed a similar trend at the age of 21 days with an average daily growth of ducks from 101.4 g to 107.6 g. The experimental group III after 21 days exceeded the control group by 7.6%.
On day 21, the body weight of ducks receiving probiotics, mannan-rich fractions, and their mix exceeded that in the control group by 1.1, 1.9 and 3.6%, respectively. The body weight was 1273 ± 67 g, 1283 ± 42 g, and 1305 ± 34 g, respectively.
In 49 days, the body weight of the ducks receiving mannan-rich fractions, as well as their mix was 3415 ± 95.5, 3459 ± 87.4, and 3547 ± 24.3 g, respectively, which exceeded the weight of the ducks of the control group by 3.1, 4.4 and 7.1 % (Table 4). In addition, a similar trend was detected with average daily gain in duck weight. In 49 days, it was 59.2, 59.7, and 61.3 g for experimental groups, exceeding that in the control group by 1.2, 2.1, and 4.7 %, respectively. The ducks receiving the mix of probiotics and mannan-rich fractions gained weight more intensively compared to the birds having the other diets (Table 5).
ВЫВОДЫ
In vitro studies showed the ability of prebiotic mannan-rich fractions isolated from the cell walls of Saccharomyces cerevisiae yeast to adsorb type I fimbriae bacterial pathogens (S. enteritidis and C. jejuni) and prevent colonization and proliferation of pathogenic microorganisms on the surface of ducks’ intestinal epithelial cells.
We studied the influence of fractions rich in mannan oligosaccharides on activity, colonization, and species composition of duck gut microflora. S. enteritidis reisolation rate decreased by 53.6% and C. jejuni ‒ by 66.2% in ducks receiving fractions rich in mannanooligosaccharides, compared to the control group. Experiments showed that the addition of prebiotic fractions to the diet did not affect the concentration of lactobacilli, bifidobacteria, enterococci, and anaerobic bacteria.
Bifido- and lactobacteria have antagonistic activity against circulating strains of S. enteritidis and C. jejuni. 88.9% of S. enteritidis isolates were susceptible to bifidobacteria and 81.5% of the studied strains were susceptible to lactobacilli. 92.6% of the isolated Campylobacter jejuni were susceptible to bifidobacteria, 76.9% of Campylbacter strains were susceptible to lactobacteria.
We developed a method of preventing bacterial infections and increasing ducks’ productivity based on the combined use of bifido- and lactobacteria (1.5×109 CFU/mL) in a ratio of 1:1 with water and fractions enriched with mannan oligosaccharides (0.4 kg/t) together with feed. We recommend the preparation from the first day of birds’ life till the end of growing period.
Preventive measures improved the preservation of the duck population by 8.76%, ensuring the average daily increase by 6.9% and the reduction of feed costs by 4.98% for 1 kg of growth throughout the growing period. During the experiment, we recorded a significant decrease in Salmonella and Campylobacter colonization in the poultry intestines and improved average daily growth. The biologically active supplements provided a significant advantage in industrial duck farming.
We demonstrated the effectiveness of natural and environmentally safe methods: yeast fractions rich in mannan oligosaccharides, probiotics, and their combined use. The method was effectively implemented in Ukrainian poultry farms.
КОНФЛИКТ ИНТЕРЕСОВ
The authors have no conflict of interest.БЛАГОДАРНОСТИ
The authors thank the managers of the poultry farms for their assistance in conducting experiments.СПИСОК ЛИТЕРАТУРЫ
- Kittler S, Fischer S, Abdulmawjood A, Glunder G, Klein G. Colonisation of a phage susceptible Campylobacter jejuni population in two phage positive broiler flocks. PLoS One. 2014;9(4). DOI: https://doi.org/10.1371/journal.pone.0094782.
- The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks breaks in 2015. EFSA Journal. 2016;14(12). DOI: https://doi.org/10.2903/j.efsa.2016.4634.
- Kasyanenko OI, Fotina TI, Fotina AA, Gladchenko SM, Gnidenko TY, Bezruk RV. Properties of Сampylobacter jejuni that were isolated from poultry products. Microbiological Journal. 2017;79(4):66–74. (In Russ.).
- Umaraw P, Prajapati A, Verma AK, Pathak V, Singh VP. Control of campylobacter in poultry industry from farm to poultry processing unit: A review. Critical Reviews in Food Science and Nutrition. 2017;57(4):659–665.
- Collard JM, Bertrand S, Dierick K, Godard C, Wildemauwe C, Vermeersch K, et al. Drastic decrease of Salmonella Enteritidis isolated from humans in Belgium in 2005, shift in phage types and influence on foodborne outbreaks. Epidemiology and Infection. 2008;136(6):771–781. DOI: https://doi.org/10.1017/s095026880700920x.
- The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA Journal. 2015;13(12). DOI: https://doi.org/10.2903/j.efsa.2015.4329.
- Meunier M, Guyard-Nicodeme M, Dory D, Chemaly M. Control strategies against Campylobacter at the poultry production level: biosecurity measures, feed additives and vaccination. Journal of Applied Microbiology. 2016;120(5):1139–1173. DOI: https://doi.org/10.1111/jam.12986.
- Skarp CPA, Hanninen ML, Rautelini HIK. Campylobacteriosis: the role of poultry meat. Clinical Microbiology and Infection. 2016;22(2):103–109. DOI: https://doi.org/10.1016/j.cmi.2015.11.019.
- Dogan ANC, Celik E, Kilicle PA, Atalay E, Saglam AG, Dogan A, et al. Antibacterial effect of hot peppers (Capsicum annuum, Capsicum annuum var globriusculum, Capsicum frutescens) on some Arcobacter, Campylobacter and Helicobacter species. Pakistan Veterinary Journal. 2018;38(3):266–270. DOI: https://doi.org/10.29261/pakvetj/2018.057.
- Spring P, Wenk C, Connolly A, Kiers A. A review of 733 published trials on Bio-Mos®, a mannan oligosaccharide, and Actigen®, a second generation mannose rich fraction, on farm and companion animals. Journal of Applied Animal Nutrition. 2015;3. DOI: https://doi.org/10.1017/jan.2015.6.
- The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA Journal. 2017;15(12). DOI: https://doi.org/10.2903/j.efsa.2017.5077.
- Ansari F, Pourjafar H, Bokaie S, Peighambari SM, Mahmoudi M, Fallah MH, et al. Association between poultry density and Salmonella infection in commercial laying flocks in Iran using a Kernel density. Pakistan Veterinary Journal. 2017;37(3):299–304.
- Gormley FJ, Bailey RA, Watson KA, McAdam J, Avendaño S, Stanley WA, et al. Campylobacter colonisation and proliferation in the broiler chicken under natural field challenge is not affected by bird growth rate or breed. Applied and Environmental Microbiology. 2014;80(21):6733–6738. DOI: https://doi.org/10.1128/AEM.02162-14.
- Ao Z, Choct M. Oligosaccharides affect performance and gut development of broiler chickens. Asian-Australasian Journal of Animal Sciences. 2013;26(1):116–121. DOI: https://doi.org/10.5713/ajas.2012.12414.
- Ramirez-Hernandez A, Rupnow J, Hutkins RW. Adherence reduction of Campylobacter jejuni and Campylobacter coli strains to HEp-2 Cells by mannan oligosaccharides and a high-molecular-weight component of cranberry extract. Journal of Food Protection. 2015;78(8):1496–1505. DOI: https://doi.org/10.4315/0362-028X.JFP-15-087.
- Muller KH, Collinson SK, Trust TJ, Kay WW. Type-1 fimbriae of Salmonella-Enteritidis. Journal of Bacteriology. 1991;173(15):4765–4772.
- Lea H, Spring P, Taylor-Pickard J, Burton E. Natural carbohydrate fraction Actigen™ from Saccharomyces cerevisiae cell wall: effects on goblet cells, gut morphology and performance of broiler chickens. Journal of Applied Animal Nutrition. 2012;1. DOI: https://doi.org/10.1017/jan.2013.6.
- Rous P, Turner JR. The preservation of living red blood cells in vitro: I. Methods of preservation. Journal of Experimental Medicine. 2016;23(2):219–237. DOI: https://doi.org/10.1084/jem.23.2.219.
- Corrigan A, de Leeuw M, Penaud-Frezet S, Dimova D, Murphy RA. Phylogenetic and functional alterations in bacterial community compositions in broiler ceca as a result of mannan oligosaccharide supplementation. Applied and Environmental Microbiology. 2015;81(10):3460–3470. DOI: https://doi.org/10.1128/aem.04194-14.
- Arsi K, Donoghue AM, Woo-Ming A, Blore PJ, Donoghue DJ. The efficacy of selected probiotic and prebiotic combinations in reducing Campylobacter colonization in broiler chickens. Journal of Applied Poultry Research. 2015;24(3):327–334. DOI: https://doi.org/10.3382/japr/pfv032.
- Hadadji M, Benama R, Saidi N, Henni DE, Kihal M. Identification of cultivable Bifidobacterium species isolated from breast-fed infants feces in West-Algeria. African Journal of Biotechnology. 2005;4(5):422–430.
- Spring P, Wenk C, Dawson KA, Newman KE. The effects of dietary mannaoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of salmonella-challenged broiler chicks. Poultry Science. 2000;79(2):205–211. DOI: https://doi.org/10.1093/ps/79.2.205.
- Pitkanen T. Review of Campylobacter spp. in drinking and environmental waters. Journal of Microbiological Methods. 2013;95(1):39–47. DOI: https://doi.org/10.1016/j.mimet.2013.06.008.
- Ugarte-Ruiz M, Florez-Cuadrado D, Wassenaar TM, Porrero MC, Dominguez L. Method comparison for enhanced recovery, isolation and qualitative detection of C. jejuni and C. coli from wastewater effluent samples. International Journal of Environmental Research and Public Health. 2015;12(3):2749–2764. DOI: https://doi.org/10.3390/ijerph120302749.
- Sujatha K, Dhanalakshmi K, Rao AS. Antigenic characterisation and antibiotic sensitivity of field isolates of Salmonella gallinarum. Indian Veterinary Journal. 2003;80(10):965–968.
- Mannan SJ, Rezwan R, Rahman MS, Begum K. Isolation and biochemical characterization of Lactobacillus species from yogurt and cheese samples in Dhaka metropolitan area. Bangladesh Pharmaceutical Journal. 2017;20(1):27–33.
- Zinedine A, Faid M. Isolation and characterization of strains of bifidobacteria with probiotic proprieties in vitro. World Journal of Dairy and Food Sciences. 2007;(1):28–34.
- Achtman M, Wain J, Weill FX, Nair S, Zhou ZM, Sangal V, et al. Multilocus Sequence Typing as a Replacement for Serotyping in Salmonella enterica. PLoS Pathogens. 2012;8(6). DOI: https://doi.org/10.1371/journal.ppat.1002776.
- Baurhoo B, Ferket PR, Zhao X. Effects of diets containing different concentrations of mannanoligosaccharide or antibiotics on growth performance, intestinal development, cecal and litter microbial populations, and carcass parameters of broilers. Poultry Science. 2009;88(11):2262–2272. DOI: https://doi.org/10.3382/ps.2008-00562.
- Rakibul Hasan AKM, Ali MH, Siddique MP, Rahman MM, Islam MA. Сlinical and laboratory diagnoses of common bacterial diseases of broiler and layer chickens. Bangladesh Journal of Veterinary Medicine. 2010;8(2):107–115. DOI: https://doi.org/10.3329/bjvm.v8i2.11188.
- Srinivasan P, Balasubramaniam GA, Murthy T, Saravanan S, Balachandran P. Prevalence and pathology of Salmonellosis in commercial layer chicken from Namakkal, India. Pakistan Veterinary Journal. 2014;34(3):324–328.
- The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA Journal. 2018;16(12). DOI: https://doi.org/10.2903/j.efsa.2018.5500.
- Safaei HG, Jalali M, Hosseini A, Narimani T, Sharifzadeh A, Raheimi E. The prevalence of bacterial contamination of table eggs from retails markets by Salmonella spp., Listeria monocytogenes, Campylobacter jejuni and Escherichia coli in Shahrekord, Iran. Jundishapur Journal of Microbiology. 2011;4(4):249–253.
- Jamshidi A, Kalidari GA, Hedayati M. Isolation and identification of Salmonella Enteritidis and Salmonella Typhimurium from the eggs of retail stores in Mashhad, Iran using conventional culture method and multiplex PCR assay. Journal of Food Safety. 2010;30(3):558–568. DOI: https://doi.org/10.1111/j.1745-4565.2010.00225.x.
- Khoult D, Krig N, Snit P. Opredelitelʹ bakteriy Berdzhi. Tom 1 [Bergey`s Manual of Determinative Bacteriology. Vol. 1]. Moscow: Mir; 1997. 432 p. (In Russ.).
