

Аннотация
Introduction. Microbial biomass is a popular source of food ingredients and feed additives. Its high use has made it focus of many relevant studies. Yeast and fungal biomasses proved to be useful substrates that improve the quality and biological value of functional products. They differ in the content and composition of proteins and polysaccharides. The present research dealt with the enzymatic decomposition of proteins found in a novel fungal and yeast biomass. The research objective was to describe the peptide and amino acid composition of their enzymatic hydrolysates.Study objects and methods. The research featured a new fungal and yeast biomass mix. Aspergillus oryzae is a mycelial fungus and a popular industrial producer of hydrolytic enzymes in food industry. As for the yeast, it was the Saccharomyces cerevisiae strain, which is often used in baking.
Results and discussion. The total content of identified amino acids in the fungal and yeast biomass was 306.0 mg/g, which was 1.5 times higher than in the fungal biomass alone. The biomass mix demonstrated a higher biological value of proteins than the yeast biomass. A set of experiments made it possible to compile a scheme for the biocatalytic destruction of polymers in the fungal and yeast biomass under the effect of fungal intracellular and endogenous enzymes. The article also contains a thorough description of the obtained enzymatic hydrolysates with various fractional compositions of peptides and free amino acids. Peptides with the molecular weight in the range of up to 29.0 kDa decreased by 2.1 times after 5 h of hydrolysis and by 10.7 times after 18 h. The designed conditions doubled the release of amino acids and increased the content of low-molecular-weight peptides up to 75.3%.
Conclusion. The research provided a new algorithm for the biocatalytic conversion of microbial biomass. Regulating the conditions of enzymatic hydrolysis made it possible to obtain enzymatic hydrolysates with a desired degree of protein degradation. They could serve as peptides and amino acids in functional food and feed products.
Ключевые слова
Microbial biomass, yeast, biocatalytic hydrolysis, enzymes, enzymatic hydrolysates, amino acids, molecular weight, peptide fractionsВВЕДЕНИЕ
The modern concept of healthy diet means that the range of functional foods keeps expanding to satisfy various physiological needs of the human organism. There are many ways to balance the nutritional and biological value of functional products, e.g. new formulations, specific raw materials, optimal technological processes, functional and biologically active additives, etc. [1–3].
The biotechnological processing of microbial biomass proved to be a promising direction for the production of functional food and feed ingredients [4–6]. Microbial biomass is a source of protein substances, vitamins, polysaccharides, and trace elements. Bacterial cell walls contain many valuable polysaccharides, including β-glucans, mannans, aminopolysaccharides, etc. [7–9]. Fungal biomass is known to produce chitosanglucan biologics [10–12]. Certain components of cell walls possess sorption, antioxidant, and other valuable properties, which makes it possible to use them in food industry [13–16]. In addition, microbial cell protoplasm contains a biologically complete protein with the amino acid score approaching that of animal protein [4, 6]. However, commercial use of microorganisms in protein and amino acid production still requires further research.
The Saccharomyces cerevisiae strain of yeast has long been focus of scientific attention. Biotechnology employs it as a substrate for protein food and feed additives. Enzyme systems can increase the bioavailability of cellular contents. They catalyze the hydrolysis of subcellular structures and release biologically valuable components, e.g. proteins [6, 17, 18]. The functional and biomedical properties of enzymatic hydrolysates depend on the degree of biocatalytic decomposition of intracellular proteins. Proteolytic enzymes owe their regulatory role to their ability to catalyze the hydrolytic degradation of the protein by certain peptide bonds. This process results in physiologically active peptides, which, in turn, can be bioregulators of certain biological processes [18–21]. The primary structure of the peptides determines their functions. Biologically active peptides (BAP) have a low molecular weight, and their amount of amino acid residues can vary from 3 to 50 [21–24].
Aspergillus fungal mycelial biomass has also been a popular subject of scientific studies. Aspergillus oryzae produces industrially significant metabolites, e.g. enzymes, organic acids, etc. [4, 10–12, 14, 25–27]. Various studies of microbial biomass as a substrate for food and feed additives revealed differences in the amount of proteins and polysaccharides. Their structure and biochemical composition also vary, which can affect the functional properties of biological products. The biomass of A. oryzae fungus contains almost twice as little protein as the S. cerevisiae yeast. However, the fungal biomass proved a valuable source of polysaccharides [25]. A fungal and yeast biomass mix can improve the quality and biological value of functional products and is a promising direction in substrate production.
Protein substances, e.g. polypeptides, low-molecularweight peptides, and amino acids, are an important component of any balanced diet. Proteins and amino acids are responsible for the formation of all tissues in a living organism. They also play a regulatory role in metabolic processes. It is the composition and amount of key amino acids that matters. This fact proves the relevance of studies aimed at obtaining various functional ingredients of food and feed products from microbial biomass as a source of biologically complete protein.
The research objective was to study the processes of enzymatic decomposition of proteins in fungal and yeast biomass. The project also focused on the effect of peptide and amino acid composition of microbial biomass enzymatic hydrolysates on the functional properties of food and feed ingredients.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
The research was performed on the premises of the Russian Research Institute of Food Biotechnology – branch of Federal Research Center of Nutrition, Biotechnology, and Food Safety (Moscow). It featured the biomass of the Aspergillus oryzae mycelial fungus, an industrial producer of hydrolytic enzymes for the food industry, and the Sаcharomyces cerevisiae strain of baker’s yeast.
The A. oryzae fungal biomass was obtained by a 10-min centrifugation at 5000 rpm. The resulting mycelial biomass was mixed with yeast in a ratio of 1:2. It served as a substrate for the biocatalytic decomposition of intracellular polymers. After centrifugation, the filtrate of the culture fluid was used to obtain a complex enzyme preparation (CEP), which served as a source of proteinases and peptidases.
The biocatalytic decomposition of the fungal and yeast biomass happened because of the autolytic processes caused by intracellular fungal enzymes. The exogenous enzymatic systems of proteolytic (CEP) and β-glucanase (Brewzyme enzyme preparation) action were introduced to increase the polymer hydrolysis.
The enzymatic activity in the enzyme systems was measured using standard methods. The mannanase activity was determined by the degree of mannan hydrolysis under certain conditions with the formation of reducing carbohydrates. The chitinase hydrolysis was assessed according to the chitin hydrolysis. State Standard R 53974-2010 was used to evaluate the general proteolytic activity, while State Standard R 53973-2010 served to measure the β-glucanase activity.
We determined the hydrolysis of the fungal and yeast biomass mix according to the concentration of reducing substances, amine nitrogen, and amino acids during enzyme hydrolysis. The anthrone method made it possible to measure the concentration of reducing substances, while the copper method helped to define the concentration of amine nitrogen [28].
We used high-pressure exclusion chromatography to assess the mass distribution of peptide molecules in the enzymatic hydrolysates. The superose 12 column (1.0 × 30 cm) was calibrated with standard globular water-soluble proteins provided by SERVA (Germany) [29]. 0.2 M sodium chloride served as an eluent at elution rate = 0 .4 cm3/min and a wavelength λ = 280 nm using a UV132 flow-through ultraviolet detector and a Multichrom 3.1 data processing software. The chromatograms were integrated by the gravimetric method. The range of molecular weights varied from free to full volume of the chromatographic column.
The research employed a KNAUER EUROCHROM 2000 chromatograph to measure the amino acid content in the microbial biomass and enzymatic hydrolysates. After that, the components were determined by a spectrophotometric Smartline UV Detector 2500 at a wavelength of λ = 570 nm (Germany). The aminograms were calculated by comparing the areas of the standard and the sample [30].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
The microbial biomass proved to vary in the amino acid composition (Table 1). The content of essential amino acids amounted to 53.13% of the total number in the fungal protein, while it was only 41.30% in the yeast protein. The level of tryptophan and methionine in the fungal protein was 2.2 times higher, leucine and tyrosine – by 1.8 times, and valine – by 1.5 times. As for the yeast protein, it appeared to contain proline; the amount of glutamic acid was by 1.4 times higer, lysine and threonine – by 1.2 times.
The yeast and fungal biomass mix had a total amino acid content of 306.0 mg/g, which was 1.5 times higher than that in the fungal biomass (202.8 mg/g). The yeast and fungal biomass had a slightly higher biological value of proteins, while the share of essential amino acids was 44.26% (Table 1).
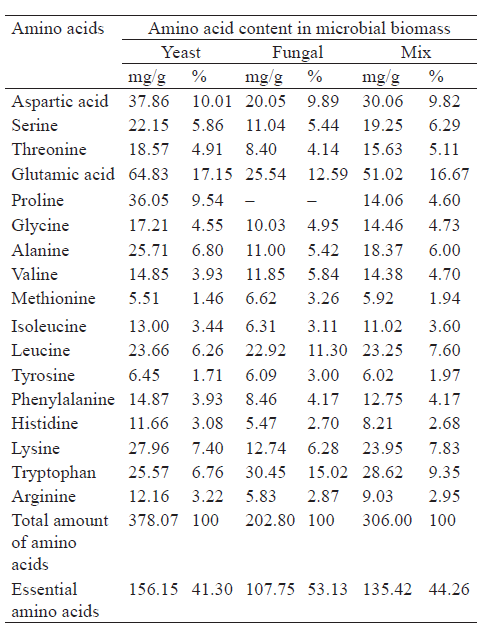
We conducted a comparative analysis of the amino acid composition of the protein in the yeast and fungal biomass mix with that of the reference protein approved by the Food and Agricultural Organization (WHO). The reference protein shows to what degree a certain protein satisfies the physiological need of the body for essential amino acids [31].
The amino acid score (ACS) was calculated according to the formula:
![]()
where ACS – amino acid score; А – essential amino acid content in a particular protein; S – amino acid content in the reference protein.
The yeast and fungal biomass demonstrated a high biological value of the protein: the total content of essential amino acids was 1.2 times higher than in the reference protein. The biomass contained two limiting amino acids, namely phenylalanine and methionine. Their amino acid score was 70% and 55% of the reference protein, respectively (Fig. 1). Tryptophan, lysine, threonine, and leucine proved to have the highest amino acid score.
Therefore, the biomass fortified with essential amino acids obtained from proteins of the S. cerevisiae yeast strain and the A. oryzae fungus can be a promising substrate for the production of new biologically active peptide and amino acid additives with a wide range of functional properties.
The microbial biomass mix had a higher level of protein in the substrate, and its biological value also increased. In addition, it demonstrated a higher content of chitin-glucan and mannan polysaccharides, as well as intracellular enzymes.
The biocatalytic conversion made it possible to increase the bioavailability of polymers in the microbial biomass mix and to obtain easily digestible peptides and amino acids. The biocatalytic conversion included three stages (Fig. 1). Stage I featured the fungal biomass, which contained residual proteolytic and β-glucanase enzymes (Table 2). The autolytic decomposition of the microbial biomass polymers lasted 2 h at 50°C. Enzymatic hydrolysate I of the biomass mix appeared after 2 h of autolysis under the effect of fungal intracellular enzymes.
The Brewzyme BGX enzyme preparation is known as a source of β-glucanases and other hydrolases (Table 2). The Brewzyme enzyme made it possible to increase the decomposition rate of cell walls during Stage II. Mannans and β-glucans, as well as protein-mannan and chitin-glucan complexes, were the main structural polymers [8, 9]. The proportion was 50 units of β-glucanase per 1 g of biomass dry matters. Stage II lasted 3 h at 40°С and produced enzymatic hydrolysate II after 5 h of hydrolysis (Fig. 2).
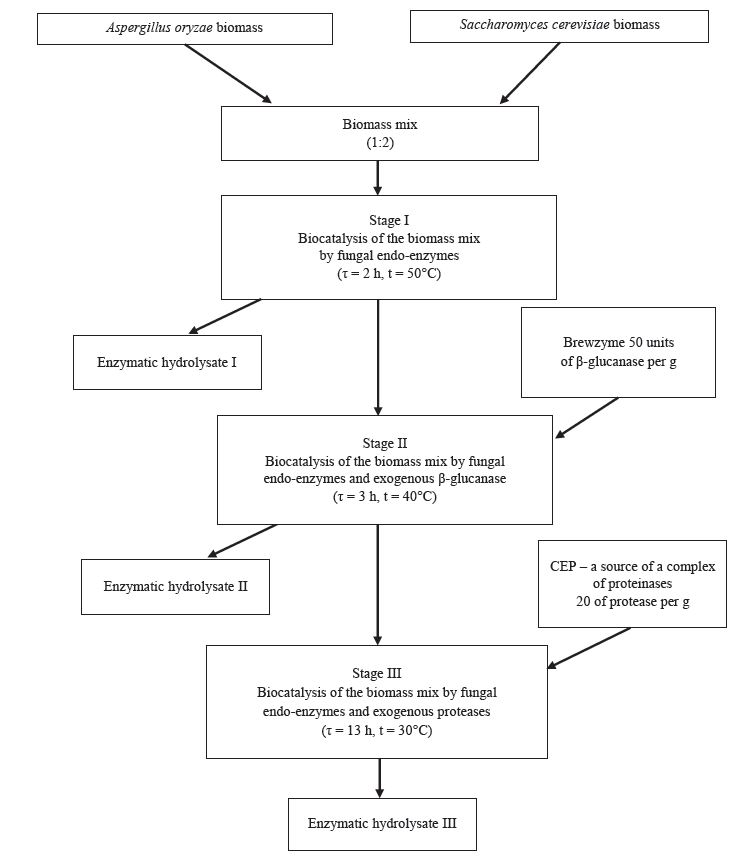
Complex enzyme preparation CEP was introduced during Stage III. It provided a deeper enzymatic hydrolysis of the main subcellular polymers of the microbial biomass, including protein substances. The hydrolysis resulted in the formation of easily digestible biologically active products. The CEP served as a source of a complex of proteinases and peptidases. The proportion was 20 units of protease per 1 g of biomass solids (Fig. 2). Fungal proteolytic enzymes are thermolabile, so the temperature was reduced to 30°C. Stage III lasted 13 h; the total biocatalysis time was 18 h. Stage III produced enzymatic hydrolysate III.
The enzyme system of the A. oryzae fungus and exogenous enzymes made it possible to obtain enzymatic hydrolysates from the yeast and fungal microbial biomass mix. The enzymatic hydrolysates varied in the degree of decomposition of intracellular polymers (Fig. 2).
The most intense formation of hydrolysis products of protein and carbohydrate polymers took place during the first 5 h. After 5 and 14 h, the concentration of soluble reducing carbohydrates increased by 9.3 and 12.1 times (from 2.1% to 25.5%), respectively. The concentration of amine nitrogen (NH2+) increased by 6.4 times and 9.6 times (from 0.5% to 4.8%). The concentration of free amino acids increased by 8.0 times and 12.2 times, from 1.3% to 15.9% (Fig. 3).
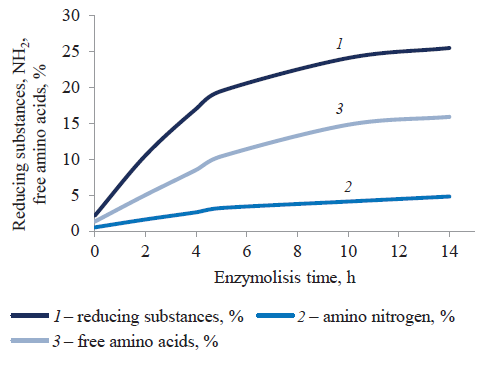
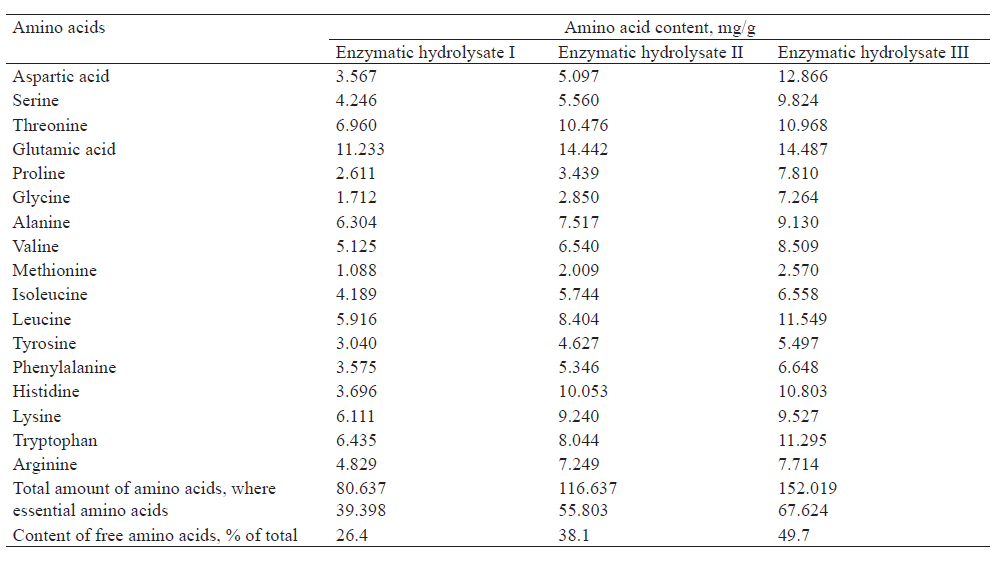
Table 3 illustrates the composition of the free amino acids in the obtained enzymatic hydrolysates and their amount. 26.4% of free amino acids were released during the hydrolysis of the microbial biomass mix by intracellular fungal enzymes (enzymatic ydrolysate I). After exogenous enzymes (β-glucanase and proteolytic effects) were introduced and the process time was prolonged, the release of amino acids increased by 1.5–2.0 times. It reached 38.1% in enzymatic hydrolysate II and 49.7% in enzymatic hydrolysate III. The content of free essential amino acids also increased (Table 3). The amount of essential free amino acids increased by 1.7 times in enzymatic hydrolysate III, rising from 39.398 mg/g to 67.624 mg/g, compared to enzymatic hydrolysate I.
Thus, controlling the enzymatic hydrolysis of the combined microbial biomass allowed us to obtain enzymatic hydrolysates with the desired degree of decomposition of microbial cell proteins.
We also measured the effect of process time on the molecular weight of the peptide fractions in the enzymatic hydrolysates of the biomass mix. Average yeast proteins consist of 466 amino acid residues and have a molecular weight of 53 kDa. Fungal roteases reduced the molecular weight of proteins after 2 h of autolysis (Fig. 4).
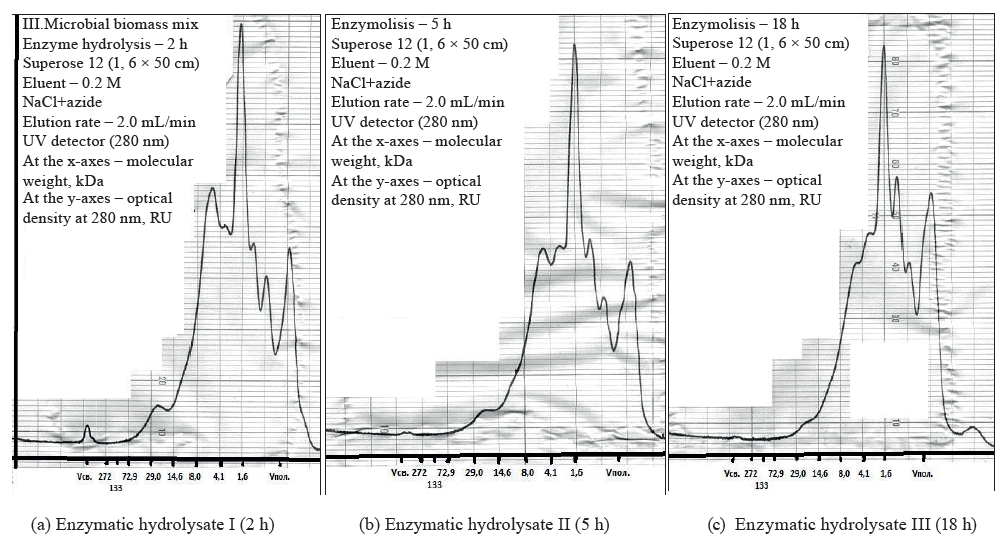
The molecular weight distribution of protein fractions during the hydrolysis of the biomass mix confirmed the effectiveness of the decomposition processes that produced lower-molecular-weight peptides (Figs. 4, 5). A longer enzymatic hydrolysis lowered the content of high-molecular-weight peptides and increased the number of low-molecular-weight peptides.
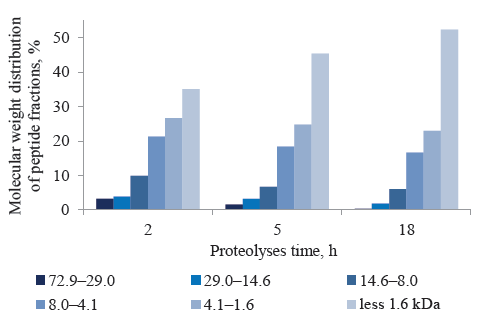
In enzymatic hydrolysate I, peptides in the range over 4.1 kDa accounted for 38.2% of the total amount of protein substances. Peptides in the range from 4.1 to 1.6 kDa constituted 26.7%, while those under 1.6 kDa made up 35.1%. The content of high-molecular-weight peptides decreased significantly during the hydrolysis of protein polymers. After 5 h of hydrolysis, the amount of peptides over 29.0 kDa fell by 2.1 times, after 18 h – by 10.7 times (Fig. 5). In enzymatic hydrolysate II, the fraction of low-molecular-weight peptides reached 45.4%. As for enzymatic hydrolysate III, the content of low-molecular-weight peptides in the range up to 4.1 kDa was 75.3%, while the share of those under 1.6 kDa accounted for 52.4%.
ВЫВОДЫ
The present research revealed the composition of peptides and amino acids in the enzymatic hydrolysates of a new biomass mix that combined the Saccharomyces cerevisiae yeast strain and the Aspergillus oryzae fungus. A set of experiments confirmed that the enzymatic hydrolysates could be used to fortify food and feed products.
The new biomass mix demonstrated a higher content of proteins and essential amino acids, as well as other valuable components. The yeast increased the amount of proteins, while the fungus raised the content of essential amino acids. The fungus also increased the amount of intracellular enzymes, which are used during enzymolisis. As a result of the mutual fortification, the total amino acid content increased by 1.5 times due to the higher protein content in the yeast. In addition, the biological value of the proteins in the new biomass mix proved to be higher than that in the traditional yeast biomass. This fact means that the ingredients obtained from the biomass mix could contribute to a wider range of functional properties.
The comparative analysis showed the high biological value of the protein in the yeast and fungal biomass mix. The total content of essential amino acids was 1.2 times higher than in the reference protein. The biomass appeared to contain two limiting amino acids – phenylalanine and methionine. Their amino acid score accounted for 70% and 55% of their content in the reference protein, respectively. Tryptophan, lysine, threonine, and leucine demonstrated the highest score. A significant amount of tryptophan, typical for fungal biomass, might add extra functional properties to ingredients obtained from their peptides and amino acids. Tryptophan is known as an immunologically active amino acid. It is a dipeptide with a wide range of immunomodulatory effects [32, 33]. Tryptophancontaining drugs have an antidepressant effect and stimulate the production of vitamin B3 (niacin). In addition, tryptophan hydroxylation produces serotonin, an important brain neurotransmitter [34].
The biomass mix fortified with essential amino acids of proteins obtained from the S. cerevisiae yeast strain and the A. oryzae fungus could be used as a commercial substrate. It was found capable of facilitating the production of new biologically active peptide and amino acid additives with a wide range of functional properties.
We developed a new algorithm for biocatalytic polymer conversion in the new microbial biomass mix. The algorithm made it possible to obtain easily digestible peptide and amino acid ingredients using fungal intracellular enzymes, as well as β-glucanase and proteolytic enzymatic preparations. The conditions of enzymatic hydrolysis proved to affect the fractional composition of the enzymatic hydrolysates. A fivehour hydrolysis lowered the amount of peptides in the range over 29.0 kDa by 2.1 times, and 18-h hydrolysis – by 10.7 times. Intracellular proteinases and peptidases are known to catalyze the decomposition of proteins. As a result, the enzymatic system with proteinases and peptidases could provide food and feed ingredients that contained 75.3% of low-molecular-weight peptides and up to 50% of free amino acids that are responsible for biologically active factors with functional properties.
The low-molecular-weight peptides, free amino acids, and essential amino acids are involved into various biological processes. They improved the digestibility of the enzymatic hydrolysates obtained from the microbial biomass mix, which can be used as peptide and amino acid components of functional food and feed products.
КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that there is no conflict of interests related to the publication of this article.ФИНАНСИРОВАНИЕ
The research was financed by the Russian Academy of Sciences (RAS) as part of a state grant within the framework of the Basic Research Program of State Academies for 2013–2020 (research topic No. 0529-2019-0066).СПИСОК ЛИТЕРАТУРЫ
- Tutelyan VA, Sukhanov BP. Food supplements: modern approaches to quality and safety. Problems of Nutrition. 2008;77(4):1–16. (In Russ.).
- Ryazanova OA, Pirogova OO. Using biologically active additives in feed supply of population. Food Industry. 2011;(2):8–10. (In Russ.).
- Gammel IV, Suvorova OV, Zaporozhskaya LI. The analysis of trends at Russian market of biologically active food supplements. Medical Almanac. 2017;51(6):154–158. (In Russ.).
- Rimareva LV, Krivova AYu, Serba EM, Overchenko MB, Ignatova NI, Pogorzelskaya NS, et al. Biological preparation based on yeast and fungal biomass rich in polysaccharides and essential amino acids. Izvestiya Ufimskogo nauchnogo tsentra RAN. 2018;(3–3):28–33. (In Russ.).
- Dhillon GS, Kaur S, Brar SK, Verma M. Green synthesis approach: extraction of chitosan from fungus mycelia. Critical Reviews in Biotechnology. 2013;33(4):379–403. DOI: https://doi.org/10.3109/07388551.2012.717217.
- Serba EM, Rimareva LV, Kurbatova EI, Volkova GS, Polyakov VA, Varlamov VP. The study of the process of enzymatic hydrolysis of yeast biomass to generate food ingredients with the specified fractional composition of protein substances. Problems of Nutrition. 2017;86(2):76–83. (In Russ.).
- Bowman SM, Free SJ. The structure and synthesis of the fungal cell wall. BioEssays. 2006;28(8):799–808. DOI: https://doi.org/10.1002/bies.20441.
- Feofilova EP. The fungal cell wall: Modern concepts of its composition and biological function. Microbiology. 2010;79(6):723–733. (In Russ.).
- Nwe N, Stevens WF, Tokura S, Tamura H. Characterization of chitosan and chitosan-glucan complex extracted from the cell wall of fungus Gongronella butleri USDB 0201 by enzymatic method. Enzyme and Microbial Technology. 2008;42(3):242–251. DOI: https://doi.org/10.1016/j.enzmictec.2007.10.001.
- Novinyuk LV, Kulev DH, Velinzon PZ, Sharova NJu. Isolation of сhitin and chitosan glucan biopolymers from mycelial waste citric acid production. Food Industry. 2016;(11):30–31. (In Russ.).
- Sharova NYu, Manzhieva BS, Printseva АА, Vybornova TV. Beta-glucans from biomass of plant and microbial origin. Food systems. 2019;2(1):23–26. DOI: https://doi.org/10.21323/2618-9771-2019-2-1-23-26.
- Kumaresapillai N, Basha RA, Sathish R. Production and evaluation of chitosan from aspergillus niger MTCC strains. Iranian Journal of Pharmaceutical Research. 2011;10(3):553–557. DOI: https://doi.org/10.22037/IJPR.2011.1003.
- Friedman M, Juneja VK. Review of antimicrobial and antioxidative activities of chitosans in food. Journal of Food Protection. 2010;73(9):1737–1761. DOI: https://doi.org/10.4315/0362-028X-73.9.1737.
- Novinyuk LV, Velinzon PZ, Kulev DKh. Sorption properties of chitinand chitosan-glucan bio-complexes isolated from Aspergillus niger fungal mycelia biomass. Proceedings of Universities. Applied Chemistry and Biotechnology. 2017;7(2)(21):64–71. (In Russ.). DOI: https://doi.org/10.21285/2227-2925-2017-7-2-64-71.
- Alsaggaf MS, Moussa SH, Tayel AA. Application of fungal chitosan incorporated with pomegranate peel extract as edible coating for microbiological, chemical and sensorial quality enhancement of Nile tilapia fillets. 2017;99:499–505. DOI: https://doi.org/10.1016/j.ijbiomac.2017.03.017.
- Tayel AA. Microbial chitosan as a biopreservative for fish sausages. International Journal of Biological Macromolecules. 2016;93:41–46. DOI: https://doi.org/10.1016/j.ijbiomac.2016.08.061.
- Sereda AS, Velikoretskaya IA, Osipov DO, Matys VYu, Bubnova TV, Nemashkalov VA, et al. The enzyme complexes for the destruction of the cell wall of filamentous fungi – producers of industrial enzymes. Izvestiya Ufimskogo nauchnogo tsentra RAN. 2018;(3–2):31–35. (In Russ.).
- Serba YeM, Overchenko MB, Pogorzhelskaya NS, Kurbatova YeI, Polyakov VA, Rimareva LV. Dependence of destruction degree in protein substances of microbe biomass on composition of proteolytic complex. Vestnik of the Russian agricultural sciences. 2015;(2):48–51. (In Russ.).
- Orlova EV, Rimareva LV, Overchenko MB, Orlova VS, Serba EM. Vliyanie fermentolizatov drozhzhey Saccharomyces cerevisiae na kletochnyy tsikl i apoptoz kletok perevivaemykh opukholey [Effect of Saccharomyces cerevisiae yeast enzymatic hydrolysates on the cell cycle and apoptosis of transplanted tumor cells]. Biozashchita i Biobezopasnost’ [Biosafety and Biosafety]. 2012;4(3)(12):48–51. (In Russ.).
- Rawlings ND, Barrett AJ, Finn R. Twenty years of the MEROPS database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Research. 2016;44(D1):D343–D350. DOI: https://doi.org/10.1093/nar/gkv1118.
- Lysenko LA, Nemova NN, Kantserova NP. Proteoliticheskaya regulyatsiya biologicheskikh protsessov [Proteolytic regulation of biological processes]. Petrozavodsk: KarRC RAS; 2011. 478 p. (In Russ.).
- Skata R. Bioaktivnye peptidy i probiotiki dlya funktsionalʹnykh myasnykh produktov [Bioactive peptides and probiotics in functional meat products]. Meat Technology. 2017;170(2):40–43. (In Russ.).
- Prosekov AYu. Osobennosti polucheniya biologicheski aktivnykh peptidov iz belkov molochnoy syvorotki [Specifics of obtaining biologically active peptides from whey proteins]. Milk Processing. 2010;127(5):12–13. (In Russ.).
- Kulikova OG, Mal’tsev DI, Il’ina AP, Burdina AV, Yamskova VP, Yamskov IA. Biologically active peptides isolated from dill Anethum graveolens L. Applied Biochemistry and Microbiology. 2015;51(3):362–366. DOI: https://doi.org/10.1134/S0003683815030114.
- Serba YeM, Rimareva LV, Overchenko MB, Sokolova YeN, Pogorzhelskaya NS, Ignatova NI, et al. Mycelia fungi – promising source of hydrolasas and valuable polymers. Vestnik of the Russian agricultural sciences. 2016;(4):41–43. (In Russ.).
- Abdel-Gawad KM, Hifney AF, Fawzy MA, Gomaa M. Technology optimization of chitosan production from Aspergillus niger biomass and its functional activities. Food Hydrocolloids. 2017;63:593–601. DOI: https://doi.org/10.1016/j.foodhyd.2016.10.001.
- Klishanets A, Luhin V, Litviak U, Trotskaya T. The chitin-glucan complex: Preparation and properties. Science and Innovations. 2016;163(9):62–67. (In Russ.).
- Instruktsiya po tekhno-khimicheskomu i mikrobiologicheskomu kontrolyu spirtovogo proizvodstva [Procedures for techno-chemical and microbiological control of alcohol production]. Moscow: DeLiprint; 2007. 479 p. (In Russ.).
- Zarin SN, Baiargargal M. Preparation of food proteins enzymatic hydrolysates of dietary proteins using some commercial enzyme preparations and various schemes of hydrolysis. Biomeditsinskaya Khimiya. 2009;55(1);73–80. (In Russ.).
- Roslyakov VYa, Tarasenko IS, Balabanov NP, Vasilʹev PS. Opredelenie kolichestva aminokislot i peptidov v preparatakh parenteralʹnogo pitaniya na osnove gidroliza belka [Determination of the amount of amino acids and peptides in parenteral nutrition preparations based on protein hydrolysis]. Russian Journal of Hematology and Transfusiology. 1984;29(3):50–52. (In Russ.).
- Yushkov S. Razrabotka kompleksnogo sostava rastitelʹnykh belkov, imeyushchego polnotsennyy nabor aminokislot [Development of a complex composition of plant proteins with a complete set of amino acids]. Biznes pishchevykh ingredientov [Business of Food Ingredients]. 2018;(1):22–27. (In Russ.).
- Faizulloeva MM, Bobizoda GM. Study of complex formation of triptophane and dipeptide of isolaicle-triptophane with zinc ion by metric titration method. News of the Academy of Sciences of the Republic of Tajikistan. Department of Biological and Medical Sciences. 2016;195(4):32–37. (In Russ.).
- Bobiev GM, Bunyatyan ND, Sayadyan KhS. Immunoaktivnye peptidy i ikh koordinatsionnye soedineniya v meditsine [Immunoactive peptides and their coordination compounds in medicine]. Moscow: Russkiy vrach; 2009. 227 p. (In Russ.).
- Pishchugin FV, Tuleberdiev IT. Kinetics and mechanism of the condensation of pyridoxal hydrochloride with L-tryptophan and D-tryptophan, and the chemical transformation of their products. Russian Journal of Physical Chemistry. 2017;91(10):1648–1652. (In Russ.). DOI: https://doi.org/10.7868/S0044453717100326.
