

Аннотация
Introduction. Various cultures of microorganisms have recently been used to accelerate technological processes. In this regard, it appears highly relevant to study the action of beneficial microorganisms on the components of food systems.Study objects and methods. The study objects included a model mixture of beef muscle and pork fat tissue with 2% salt, as well as a model protein. Lactobacillus plantarum and Staphylococcus carnosus were used in an amount of 1×107 CFU/g of raw material. The compositions of free amino and fatty acids, carbohydrates, and other components were analyzed by liquid and gas chromatography with mass-selective detection.
Results and discussion. We studied the effect of L. plantarum and S. carnosus on protein, lipid, and carbohydrate components of food systems based on animal raw materials. We found that the combined effect of the cultures was by 25% as effective as their individual use at 4×109 CFU/kg of raw material. The three-week hydrolysis of proteins to free amino acids was almost a third more effective than when the cultures were used separately. The synergistic effect of L. plantarum and S. carnosus on fat components was not detected reliably. Free monosaccharides formed more intensively when the cultures were used together. In particular, the amount of free lactose almost doubled, compared to the cultures’ individual action.
Conclusion. We described culture-caused quantitative changes in the main components of animal-based food systems: amino acids, fatty acids, carbohydrates, and basic organic compounds. Also, we identified substances that can affect the taste and aroma of final products when the cultures are used together or separately. These results make it possible to obtain products with a wide variety of sensory properties.
Ключевые слова
Sensory properties, Lactobacillus plantarum, Staphylococcus carnosus, food systems, meat products, microorganismsВВЕДЕНИЕ
Microorganisms have long been widely used in the food industry: in winemaking and beverage production, dairy production, silage making and fermentation of plant materials, as well as in seafood fermentation [1–17].
Products of animal origin make up a significant part of total food consumption. Like any products, they undergo biochemical transformations during production and are exposed to microorganisms during long-term storage [18–19].
Literature describes various methods for producing foods under the influence of microorganisms contained in animal raw materials. One of such methods – aging – is mainly used to produce dry-cured products. This process involves curing raw materials and semi-products under certain conditions to expose them to a gradual, and sometimes fairly long, effect of the microorganisms’ enzyme systems. As a result, the main food components transform and develop certain flavoring characteristics [20–22].
Today, formulators forcibly introduce starter cultures of microorganisms into raw materials to reduce process time. In particular, for food systems based on animal raw materials, they use Staphylococcus (St. xylosus, St. carnosus), Lactobacillus (L. pentosus, L. plantarum, L. sakei, L. curvatus), and Pediococcus (P. pentosaceus, P. acidilactici) [23–26]. Microorganisms can produce different effects and form a wide variety of flavors [27–28].
For convenience, microorganisms are commonly used as lyophilized solids, with culture cells deposited on the surface of a solid carrier, usually sugars. For products of animal origin, 1–2 g of a freeze-dried culture containing (1–10)×1012 CFU/100 g sucrose is usually added per every 5 or 10 kg of raw material [21, 29].
Although the action of microbiological cultures on various food materials is fully described, scientific literature lacks systematic data on the quantitative changes in the most important minor components of animal materials (individual amino acids, fatty acids, and monosugars) under the influence of L. plantarum and S. carnosus.
We aimed to identify the effects of microorganism cultures on protein, lipid, and carbohydrate components of animal-based food systems.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Our objects of the study included a model food system – a mixture of beef muscle tissue Longissimus dorsi and pork fat tissue Telae adipem with 2% sodium chloride (75:25%) homogenized in a Buchi Mixer B400 blender (Switzerland), as well as a model protein. The model food system contained 18.5% protein, 23% fat, 2.5% carbohydrates, and 54% moisture.
The model protein was obtained by a 6 h extraction at 25°C of Longissimus dorsi of Bubulae beef with a 5% sodium chloride solution followed by desalting on G25 and freeze-drying [30]. The isolated protein was 93% pure and had a 6% moisture.
Lactobacillus plantarum ATCC 8014 (LP) and Staphylococcus carnosus ATCC 51365 (SC) were added in an amount of 1×107 CFU/g of raw material. We used preparations of culture on freeze-dried sucrose in an amount of 2×1010 CFU/g.
The model mixture was treated as follows. First, animal raw materials were kept in salt for 24 h at 2 ± 2°С. Then, we introduced starter cultures and packed the mixture in plastic bags to keep in the chamber for 5 days at 2–4°С, relative humidity (W) 85%, and an air flow speed of 0.1 m/s. Further treatment was carried out during 5 days (15°C, W 82%) and 10 days (12°C, W 75%). The control sample was kept in salt for 24 h at 2 ± 2°C.
The model protein was treated with starter cultures in a 2% sodium chloride solution (hydromodule 1:5) under similar conditions at pH 7.0.
To measure the proteolytic activity, we placed a 1% casein solution in 0.05 M Tris-PO4 buffer (pH 7.0) into two tubes (5 mL in each) and added 10 mL of distilled water to the first tube and 1 mL of a 1×1010 CFU/mL enzyme solution or 1 mL of the test solution to the second tube. After a 10-min exposure at 37°C, we added 5 mL of a 10% trichloroacetic acid solution to the test samples, filtered them through a 0.45 μm filter, and measured the optical densities of the transparent solutions against the control at 280 nm. The proteolytic activity (units/mg) was calculated as A = ( D280 sample – D 280 control)/10·g, where g is the nominal enzyme concentration in the test sample. The standard unit of peptidase activity is the amount of enzyme required to release free amino acids during proteolytic decomposition. It is equivalent to a change in the absorption rate of the test solution (0.001D280) per minute at 37°C and pH 7.0 [31].
The materials were treated with L. plantarum and S. carnosus in a 1:5 ratio: 1 g of the enzyme preparation per 5 kg of the formulation and 1 mg of the preparation per 5 g of animal protein.
The content of amino acids was determined on a Biotronic 6001 amino acid analyzer (Germany) by distribution chromatography after acid hydrolysis of proteins [31].
Free amino acids were determined after protein precipitation by adding 10% trichloroacetic acid, followed by neutralization with a 10 M sodium hydroxide solution to pH 2.0 and filtration through a Millipore membrane filter with a pore diameter of 0.22 μm. Then, the filtrate was diluted in a buffer solution (pH 2.2). To quantify individual amino acids, we compared the peak areas in the aminogram obtained with the Winpeak Eppendorf-Biotronic integration system (Germany) by analyzing a standard mixture of amino acids that contains 2.5 μmol of each amino acid in 1 mL of the solution [31].
Fatty acids and chemical components responsible for the product’s taste and aroma were determined by chromatography-mass spectrometry [21, 31].
The components were analyzed on a 7890A gas chromatograph with a 5975C VLMSD mass selective detector (Agilent Technologies, USA) using a modified Folch method. In particular, a 1 g sample was subjected to a mixture of 10 mL chloroform and 10 mL methanol in the presence of a 1% KCl solution for 24 h to dissolve the lipid components. The extract was filtered through paper. After removing the excess solvents by evaporation to dryness, the residue was subjected to acid hydrolysis to obtain methyl esters of fatty acids, which were analyzed by gas chromatography.
A 0.01 g amount of lipids was treated with 3 mL of a 15% solution of acetyl chloride in methanol at 100°C for 2 h. Then, the mixture was neutralized by KOH (1.25 mL) in СН3ОН to pH 5.0–6.0. A few minutes after adding 3 mL of a saturated aqueous NaCl solution and 3 mL of hexane, we took for analysis 0.2 μL from a clear hexane layer containing methyl esters of fatty acids. Chromatography was performed on a 30 m×0.32 mm×0.5 μm HP-Innowax capillary column under the following conditions: the column temperature in the thermostat increasing from 100°C to 260°C at a rate of 10°C/min; injector temperature 250°C, detector temperature 300°C; hydrogen flow from the generator at 35 cm3/min; nitrogen flow at 20 cm3/min; flow division 1:100; analysis time 30 min; injection of 1 μL of the sample. A NIST08 MS Library was used to measure the content of isomers, an automatic search and identification program for gas chromatography-mass spectrometry with a probability of peaks correlation above 65%.
The content of free fatty acids was determined by an acid-base titration of the samples according to the acid number. In 2 mg KOH/g of the product, it corresponded to 1% of the mass content of free fatty acids [31].
The composition of free carbohydrates was analyzed using a BioLC chromatographic system, including a GS50 gradient pump, an ED50 electrochemical detector, an EG50 eluent generator with 10 mN NaOH, and an LC25 chromatographic thermostat with a CarboPac PA20 column (Dionex, Germany). The content of free carbohydrates was determined in aqueous extracts of 0.01 g of the sample in 100 mL of demineralized HPLCgrade water filtered through a 0.45 μm filter at 25°C.
The water retention capacity was determined by a standard method, recording bound moisture under load [31].
Our study used casein, tris (hydroxymethyl) aminomethane (tris), phosphoric acid, sodium chloride, sodium hydroxide, potassium hydroxide, and Sephadex G-25 (Sigma, USA). As standards of amino acids, we used a solution of mixed individual amino acids in a molar concentration of 2.5 μmol/mL (Supelco, USA): glycine, alanine, valine, leucine, isoleucine, proline, phenylalanine, tyrosine, methionine, cysteine, aspartic acid, glutamic acids, lysine, arginine, histidine, serine, and threonine.
As standards of fatty acids, we used a solution of mixed C6–C24 fatty acid methyl esters in methylene chloride with a mass concentration of 10 mg/mL (Supelco, USA): caproic (C6:0), octanoic (C8:0), decanoic (C10:0), decenoic (C10:1), undecanoic (C11:0), dodecanoic (C12:0), tridecanoic (C13:0), tetradecanoic (C14:0), cis-9-tetradecenoic (C14:1), pentadecanoic (C15:0), cis-10-pentadecenoic (C15:1), hexadecanoic (C16:0), cis-9-hexadecenoic (C16:1), heptadecanoic (C17:0), cis-10-heptadecenoic (C17:1), octadecanoic (C18:0), cis-9-octadecenoic (C18:1n9c), trans-9-octadecenoic (С18:1n9t), cis-9,12-octadecadienoic (С18:2n6), cis-6,9,12-оctadecatrienoic (С18:3n6), cis-9,12,15-оctadecatrienoic (С18:3n3), nonadecanoic (С19:0), eicosanoic (С20:0), cis-9-eicosenoic (С20:1n9), cis-11,14,17-eicosatrienoic (C20:3n3), cis-8,11,14-eicosatrienoic (C20:3n6), cis-11,14,17-eicosatrienoic (C20:3n3), cis-5,8,11,14-eicosatetraenoic (C20:4n6), eicosapentaenoic (C20:5n3), heneicosanoic (C21:0), docosanoic (C22: 0), cis-13-docosenoic (C22:1n9), cis-13,16-docosadienoic (C22:2n6), cis-7,10,13,16,19-docosapentaenoic (C22:5n3), cis-4,7,10,13,16,19-docosahexaenoic (C22:6n3), tricosanoic (C23:0), tetracosanoic (C24:0), and cis-15-tetracosenoic (C24:1).
As carbohydrate standards, we used arabinose (Ara, C5H10O5, D-(−)-arabinose ≥ 99%, A3131 Sigma), galactose (Gal, C6H12O6, D-(+)-galactose ≥ 99%, G0750 Sigma-Aldrich), glucose (Glc, C6H12O6, D-(+)-glucose ≥ 99.5%, G8270 Sigma), xylose (Xyl), mannose (Man, C6H12O6, D-(+)-mannose from wood, ≥ 99% M2069 Sigma), fructose (Fru, C6H12O6, D-(−)-fructose ≥ 99%, F0127 Sigma), sucrose (Sug, C12H22O11, α-D-glucose-(1→2)-β-D-fructose, sucrose ≥ 99.5% S9378 Sigma), ribose (C5H10O5, D-(−)-ribose ≥ 99% R7500 Sigma), lactose (Lac, C12H22O11·H2O, β-D-galactose-(1→4)-α-Dglucose, α-Lactose monohydrate reagent grade L3625 Sigma-Aldrich), aqueous solutions in a concentration of 0.001 mg/mL.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Animal-based products have a protein content of 10 to 25% [18, 29]. Fresh raw materials of animal origin usually contain from 0.001 to 0.01% of free amino acids, and their content increases with prolonged storage due to internal enzyme systems. We determined the amino acid composition of the model protein and the meat system protein before and after introducing Lactobacillus plantarum and Staphylococcus carnosus (Table 1). We found that the starter cultures significantly increased the total content of free amino acids both in the model protein and in the formulation. In the formulation, their content increased to 2.0 ± 0.1%, 2.2 ± 0.1%, and 2.8 ± 0.1% after using Lactobacillus plantarum, Staphylococcus carnosus, and an equimolar mixture of Lactobacillus plantarum and Staphylococcus carnosus, respectively.
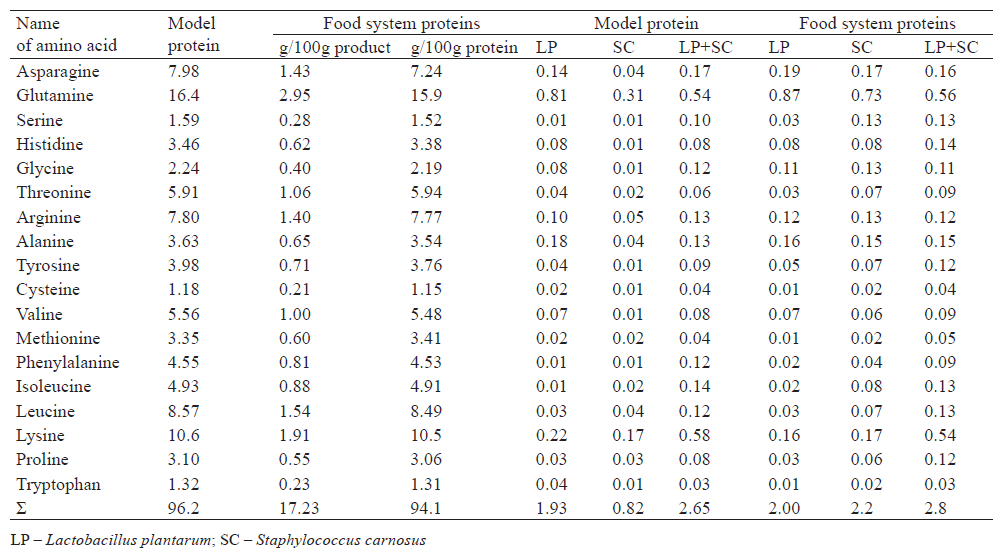
With the same total concentration of introduced cultures at 4×109 CFU/kg, the mixture of Lactobacillus plantarum and Staphylococcus carnosus increased the rate of protein hydrolysis to free amino acids by 30–40% (P > 0.96), compared to their separate action.
To determine the treatment time, we observed changes in the release of free amino acids and the correlating values of water retention (Fig. 1). We found that the optimal treatment time was three weeks. This time ensured optimal quality parameters of the product, including water retention (Fig. 1).
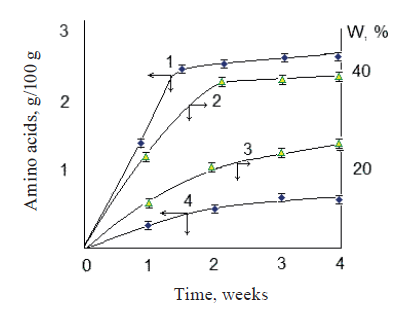
According to Fig. 1, a joint use of the bacterial cultures ensured the maximum concentration of free amino acids in two weeks. In addition, the water retention capacity of the food system – an important technological indicator of product quality – was almost twice as high as when the process lasted a month without using bacterial cultures.
The proteolytic activity with respect to the model and food system proteins showed a synergistic effect of L. plantarum and S. carnosus on the protein components (Table 2). Their joint use increased the efficiency of hydrolytic decomposition of the system proteins leading to a release of free amino acids (Table 1).

Table 3 shows the composition of fatty acids in the animal-based food system and changes in their contents under the influence of L. plantarum and S. carnosus. At the initial stages, we found a decrease in the contents of lower C6–C10 and unsaturated fatty acids, especially essential omega-3 acids (α-оctadecatrienoic C18:3, eicosapentaenoic C20:5, and docosapentaenoic C22:5). However, there was an increase in cis-11,14,17-eicosatrienoic С20:3 acid that is important for proper nutrition of mammals. Three weeks of treating the food system with L. plantarum and S. carnosus led to a decrease in unsaturated fatty acids and an increase in saturated acids by 1–5% (P > 0.95). Similar changes in contents of saturated and unsaturated fatty acids are usually observed for animal-based products subjected to long-term storage at low temperatures [22, 23].
We did not evaluate the direct lipolytic activity of L. plantarum and S. carnosus in the presence of synthetic substrates commonly used for this purpose. However, Table 3 shows that a combined action of the cultures on the system led to a more efficient breakdown of animal fats and a release of free fatty acids, compared to their individual action. Yet, this effect was not expressed clearly.
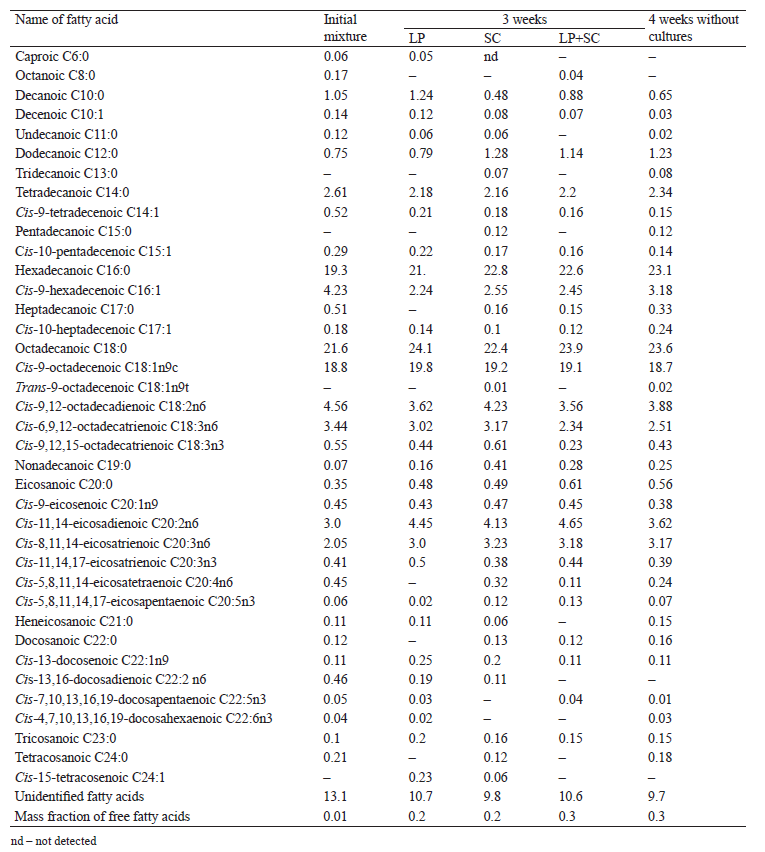
Thus, the action of L. plantarum and S. carnosus on the fat components of the food system not only transformed the fatty acid composition, but also, to a much greater extent, increased the amount of free fatty acids.
Figure 2 shows a kinetic curve of changes in the content of free fatty acids as a result of treatment with L. plantarum and S. carnosus. Longer treatment time led to a higher mass fraction of free fatty acids in all cases. We found no differences in the kinetics of free fatty acids formation under individual or joint action of the cultures, apparently due to their comparable lipolytic activity.
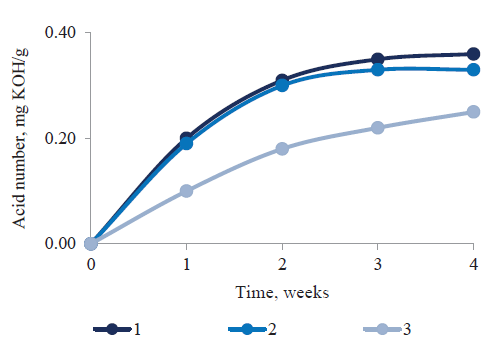
The synergistic effect of L. plantarum and S. carnosus on the fat components of the animalbased food system did not manifest reliably, since the amounts of free fatty acids in the final products were approximately the same.
Thus, the action of L. plantarum and S. carnosus resulted in not only the hydrolytic decomposition of fat components, but also in their biochemical transformation into ultimate chemical structures. The increased content of saturated acids found in our study should be considered critically, in light of current trends in the production of foods with an increased amount of unsaturated, especially polyunsaturated, fatty acids of the omega-3 family. This problem should be taken into account in further development of methods for producing animal-based foods.
Table 4 shows the quantitative identification of free carbohydrates in the food system treated with L. plantarum and S. carnosus. As we can see, the content of some sugars (galactose, glucose, fructose) decreased, while the content of others increased. It appears that the decrease was caused by the consumption of those sugars by the cultures themselves, whereas the increase was associated with proteolytic and carbohydrolase activity, leading to the breakdown of animal polysaccharides. The mass fraction of such polysaccharides in animal raw materials is 2–3%, but their decomposition can lead to the formation of 0.1–100 mg% free carbohydrates [18, 23].
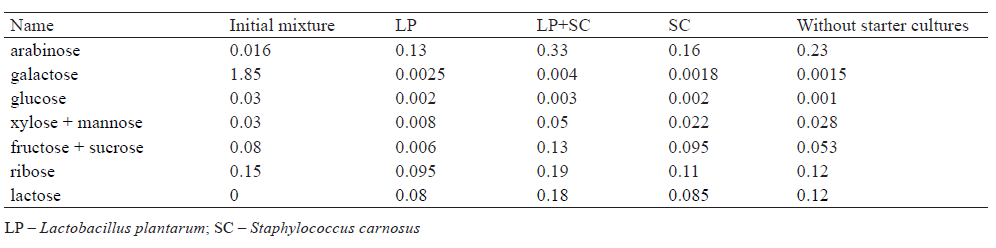
According to Table 4, free monosaccharides formed most intensively under the joint action of L. plantarum and S. carnosus. It manifested through changes in the content of galactose, glucose, xylose, and ribose and through a higher rate of disaccharides (lactose and sucrose) formation in the food system. In fact, the amount of free lactose was almost twice as high as when the cultures were used individually. As a result, the product acquired a sweetish taste. Thus, L. plantarum and S. carnosus produced a synergistic effect on the changes in carbohydrate components of the animalbased food system.
Further, treating food systems with microcultures totally changes the chemical composition of organic substances in raw materials and intermediates or those substances formed as a result. Table 5 shows changes in some minor components of the food system in the presence of L. plantarum and S. carnosus. The mass spectrometric analysis made it possible to identify over 250 organic compounds which could be considered as a result of biochemical effects that microorganisms had on protein, lipid, and carbohydrate components of the food system. A large amount of those compounds were derivatives of fatty acids. Table 5 lists the main substances identified, whose content exceeded 0.001%.
As can be seen in Table 5, the joint use of L. plantarum and S. carnosus resulted in more pronounced changes in almost all compounds, compared to their individual action. To some extent, this result indicated a synergistic mechanism of action of both cultures used to treat animal raw materials.
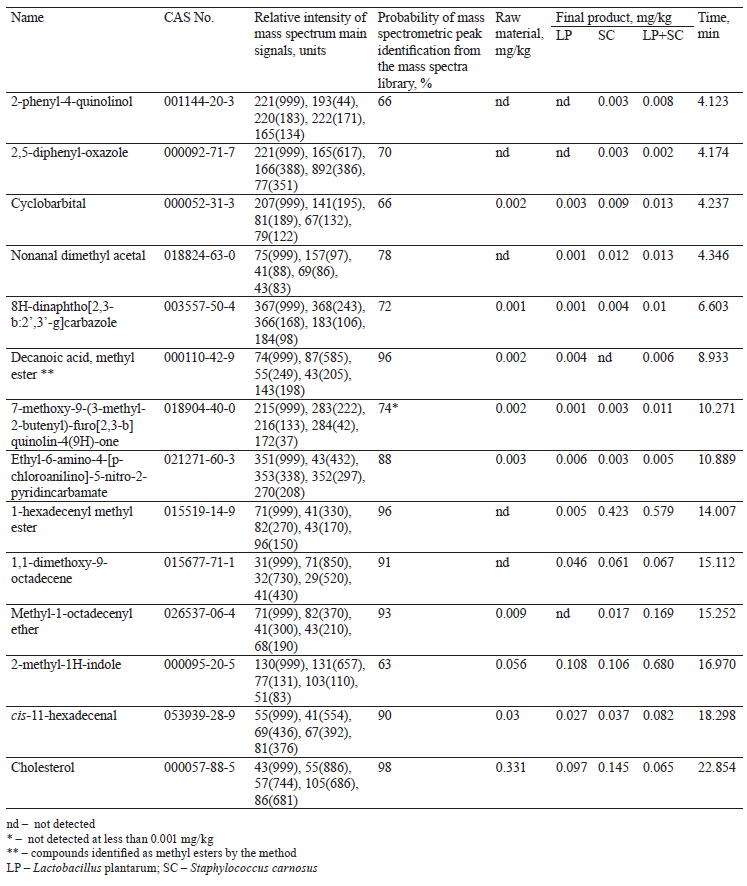
Given that all identified substances can affect the taste and aroma of final products, varying the use of starter cultures – both individual and joint – can make it possible to obtain products with a wide range of consumer properties [21].
Finally, the biochemical transformations of cholesterol require our special attention (Table 5). This substance is a significant component of food systems based on animal raw materials. Its high content in products is considered as an unfavorable factor leading to the development of atherosclerosis. In our case, the combined action of the cultures led to a more considerable degradation of cholesterol, which is an important advantage of using these cultures together.
ВЫВОДЫ
Thus, the joint use of starter cultures, Lactobacillus plantarum and Staphylococcus carnosus, to treat animal-based food systems not only increased the yield of the product, but also had a synergistic effect on the protein, lipid, and carbohydrate components of the system. This may allow us to change the component composition of the system and form the desired characteristics of the food product.КОНФЛИКТ ИНТЕРЕСОВ
The authors declare that there is no conflict of interests.СПИСОК ЛИТЕРАТУРЫ
- Takahashi M, Masaki K, Mizuno A, Goto-Yamamoto N. Modified COLD-PCR for detection of minor microorganisms in wine samples during the fermentation. Food Microbiology. 2014;39:74–80. DOI: https://doi.org/10.1016/j.fm.2013.11.009.
- Cimaglia F, Tristezza M, Saccomanno A, Rampino P, Perrotta C, Capozzi V, et al. An innovative oligonucleotide microarray to detect spoilage microorganisms in wine. Food Control. 2018;87:169–179. DOI: https://doi.org/10.1016/j.foodcont.2017.12.023.
- Longin C, Petitgonnet C, Guilloux-Benatier M, Rousseaux S, Alexandre H. Application of flow cytometry to wine microorganisms. Food Microbiology. 2017;62:221–231. DOI: https://doi.org/10.1016/j.fm.2016.10.023.
- Ribes S, Ruiz-Rico M, Pérez-Esteve E, Fuentes A, Barat JM. Enhancing the antimicrobial activity of eugenol, carvacrol and vanillin immobilised on silica supports against Escherichia coli or Zygosaccharomyces rouxii in fruit juices by their binary combinations. LWT – Food Science and Technology. 2019;113. DOI: https://doi.org/10.1016/j.lwt.2019.108326.
- Bracke N, Van Poucke M, Baert B, Wynendaele E, De Bels L, Van den Broeck W, et al. Identification of a microscopically selected microorganism in milk samples. Journal of Dairy Science. 2014;97(2):609–615. DOI: https://doi.org/10.3168/jds.2013-6932.
- Meng L, Zhang YD, Liu HM, Zhao SG, Wang JQ, Zheng N. Characterization of Pseudomonas spp. and associated proteolytic properties in raw stored at low temperatures. Frontiers in Microbiology. 20178. DOI: https://doi.org/10.3389/fmicb.2017.02158.
- Glück C, Rentschler E, Krewinkel M, Merz M, von Neubeck M, Wenning M, et al. Thermostability of peptidases secreted by microorganisms associated with raw milk. International Dairy Journal. 2016;56:186–197. DOI: https://doi.org/10.1016/j.idairyj.2016.01.025.
- Dallas DC, Citerne F, Tian T, Silva VLM, Kalanetra KM, Frese SA, et al. Peptidomic analysis reveals proteolytic activeity of kefir microorganisms on bovine milk proteins. Food Chemistry. 2016;197:273–284. DOI: https://doi.org/10.1016/j.foodchem.2015.10.116.
- Ribeiro JC, Peruzi GAS, Bruzaroski SR, Tamanini R, Lobo CMO, Alexandrino B, et al. Short communication: Effect of bactofugation of raw milk on counts and microbial diversity of psychrotrophs. Journal of Dairy Science. 2019;102(9):7794–7799. DOI: https://doi.org/10.3168/jds.2018-16148.
- Yang LL, Yang XJ, Li JF, Dong ZH, Shao T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresource Technology. 2019;275:280–287. DOI: https://doi.org/10.1016/j.biortech.2018.12.067.
- Cao P, Wu LY, Wu Z, Pan DD, Zeng XQ, Guo YX, et al. Effects of oligosaccharides on the fermentation properties of Lactobacillus plantarum. Journal of Dairy Science. 2019;102(4):2863–2872. DOI: https://doi.org/10.3168/jds.2018-15410.
- Xu YM, Cui YL, Wang X, Yue FF, Shan YY, Liu BF, et al. Purification, characterization and bioactivity of exopolysaccharides produced by Lactobacillus plantarum KX041. International Journal of Biological Macromolecules. 2019;128:480–492. DOI: https://doi.org/10.1016/j.ijbiomac.2019.01.117.
- Geeta, Yadav GA. Antioxidant and antimicrobial profile of chicken sausages prepared after fermentation of minced chicken meat with Lactobacillus plantarum and with additional dextrose and starch. LWT – Food Science and Technology. 2017;77:249–258. DOI: https://doi.org/10.1016/j.lwt.2016.11.050.
- Aret VА. Use of food resources and development of food production technology. Foods and Raw Materials. 2017;5(1):4–10. DOI: https://doi.org/10.21179/2308-4057-2017-1-4-10.
- Lazado CC, Caipang CMA, Estante EG. Prospects of host-associated microorganisms in fish and penaeids as probiotics with immunomodulatory functions. Fish and Shellfish Immunology. 2015;45(1):2–12. DOI: https://doi.org/10.1016/j.fsi.2015.02.023.
- Solli L, Schnürer A, Horn SJ. Process performance and population dynamics of ammonium tolerant microorganisms during co-digestion of fish waste and manure. Renewable Energy. 2018;125:529–536. DOI: https://doi.org/10.1016/j.renene.2018.02.123.
- Falas P, Jewell KS, Hermes N, Wick A, Ternes TA, Joss A, et al. Transformation, CO2 formation and uptake of four organic micropollutants by carrier-attached microorganisms. Water Research. 2018;141:405–416. DOI: https://doi.org/10.1016/j.watres.2018.03.040.
- Majou D, Christieans S. Mechanisms of the bactericidal effects of nitrate and nitrite in cured meats. Meat Science. 2018;145:273–284. DOI: https://doi.org/10.1016/j.meatsci.2018.06.013.
- Torngren MA, Darré M, Gunvig A, Bardenshtein A. Case studies of packaging and processing solutions to improve meat quality and safety. Meat Science. 2018;144:149–158. DOI: https://doi.org/10.1016/j.meatsci.2018.06.018.
- Janssens M, Myter N, De Vuyst L, Leroy F. Species diversity and metabolic impact of the microbiota are low in spontaneously acidified Belgian sausages with an added starter culture of Staphylococcus carnosus. Food Microbiology. 2012;29(2):167–177. DOI: https://doi.org/10.1016/j.fm.2011.07.005.
- Simonova M, Strompfova V, Marcinakova M, Laukovda A, Vesterlund S, Moratalla ML, et al. Characterization of Staphylococcus xylosus and Staphylococcus carnosus isolated from Slovak meat products. Meat Science. 2006;73(4):559–564. DOI: https://doi.org/10.1016/j.meatsci.2006.02.004.
- Ivankin AN, Oliferenko GL, Kulikovskii AV, Chernukha IM, Semenova AA, Spiridonov KI, et al. Determination of unsaturated fatty acids with a migrating double bond in complex biological matrices by gas chromatography with flame ionization and mass spectrometry detection. Journal of Analytical Chemistry. 2016;71(11):1131–1137. DOI: https://doi.org/10.1134/S1061934816110046.
- Stavropoulou DA, De Maere H, Berardo A, Janssens B, Filippou P, De Vuyst L, et al. Pervasiveness of Staphylococcus carnosus over Staphylococcus xylosus is affected by the level of acidification within a conventional meat starter culture set-up. International Journal of Food Microbiology. 2018;274:60–66. DOI: https://doi.org/10.1016/j.ijfoodmicro.2018.03.006.
- Lorenzo JM, Munekata PES, Domínguez R. Role of autochthonous starter cultures in the reduction of biogenic amines in traditional meat products. Current Opinion in Food Science. 2017;14:61–65. DOI: https://doi.org/10.1016/j.cofs.2017.01.009.
- Palmieri G, Balestrieri M, Capuano F, Proroga YTR, Pomilio F, Centorame P, et al. Bactericidal and antibiofilm activity of bactenecin-derivative peptides against the food-pathogen Listeria monocytogenes: New perspectives for food processing industry. International Journal of Food Microbiology. 2018;279:33–42. DOI: https://doi.org/10.1016/j.ijfoodmicro.2018.04.039.
- de Almeida MA, Saldana E, Pinto JSD, Palacios J, Contreras-Castillo CJ, Sentandreu MA, et al. A peptidomic approach of meat protein degradation in a low-sodium fermented sausage model using autochthonous starter cultures. Food Research International. 2018;109:368–379. DOI: https://doi.org/10.1016/j.foodres.2018.04.042.
- Manyukhin YaS, Tchernukha IM, Kovalyov LI, Ivanov AV, Kovalyova MA, Shishkin SS. The study of horsemeat proteins by use proteomic technologies. Vsyo o Myase. 2014;(3):20–25. (In Russ.).
- Zagustina NA, Misharina TA, Veprizky AA, Zhukov VG, Ruzhitsky AO, Terenina MB, et al. Elimination of volatile compounds of leaf tobacco from air emissions using biofiltration. Applied Biochemistry and Microbiology. 2012;48(4):385–395. DOI: https://doi.org/10.1134/S000368381204014x.
- Ivankin AN, Semenova AA, Nasonova VV, Kulikovskii AV, Vostrikova NL, Rogatin AI, et al. Biotechnology for formation of aromatic properties of national-food foodstuffs on the basis of meat raw material under influence of bacterial crops and chromato-mass-spectrometric analysis of the flavoring components. Journal of Applied Biotechnology and Bioengineering. 2017;3(4):366–372. DOI: https://doi.org/10.15406/jabb.2017.03.00072.
- Aaslyng MD, Vestergaard C, Koch AG. The effect of salt reduction on sensory quality and microbial growth in hotdog sausages, bacon, ham and salami. Meat Science. 2014;96(1):47–55. DOI: https://doi.org/10.1016/j.meatsci.2013.06.004.
- Lisitsyn AB, Ivankin AN, Neklyudov AD. Metody prakticheskoy biotekhnologii. Analiz komponentov i mikroprimesey v myasnykh i drugikh pishchevykh produktakh [Methods of practical biotechnology. Analysis of components and trace elements in meat and other food products]. Moscow: VNIIMP; 2002. 408 p. (In Russ.).
