

Аннотация
The effect of animal and plant proteases as well as starters, or starter cultures, on protein aggregates for- mation in raw pork and beef as well as meat products was studied. The proteomic analysis of raw meat revealed that animal proteases – pepsin and trypsin – caused the aggregation of isoform 2 of protein 1 containing 4.5 LIM domains. Vacuum packaged meat showed the same results during storage, while unpacking led to the acceleration of the aggregation process due to autolysis. In addition, mixed aggregated fragments, such as muscle creatine phos- phokinase and glutathione-S-transferase, actin and perilipin, and type II keratin appeared in those samples. Starters with Pediococcus pentosaceus 31 from the Russian National Collection of Industrial Microorganisms (VKPM-8901) caused myoglobin and troponin I aggregation, while the formation of soy proteins aggregates (glycinin G1 and glyci- nin A3B4) was detected in meat products as a result of the autolysis process and the use of cholesterol-lowering star- ters. All in all, proteases which cause protein aggregation may be less effective for raw meat tenderization, whereas the proteins identified may be used as quality biomarkers.Ключевые слова
Proteins, meat, proteases, aggregates, biomarkersВВЕДЕНИЕ
Protein aggregation in this paper is defined as the interaction of denatured protein molecules resulting in the formation of weak or strong (for example, disulfide) molecular linkages. These linkages contribute to larger particles formation. Mostly, protein aggregation in meat occurs as a result of thermal denaturation when heating or autolysis in the rigor mortis stage.
Denatured and then aggregated proteins become less soluble and lose their biological properties. Rigor mortis also causes changes which lead to protein aggregation.
On the other hand, proteolytic enzymes are capable of weakening intermolecular interactions in proteins and provide proteolysis, which contributes to tissue tenderization [1]. Both proteolytic and meat enzymes act on protein substrate fragments and thereby influence protein components structures. The enzymes facilitate aggregates dissotiation, free SH-groups formation, and muscle tissue properties partial recovery after denaturation or rigor mortis. As a result of the enzyme impact, meat becomes tender, juicy, with pronounced taste and aroma.
The proteolytic activity of lactic-acid bacteria strains against sarcoplasmic and myofibrillar proteins was studied by different research groups [2-4].
In 2015, the hydrolysis of myofibrillar proteins during the fermentation of sausage models with starters or their mixture was studied by Lopez et al. [2]. The researchers used such starters as Lactobacillus curvatus CRL705 and Staphylococcus vitulinus GV318. The results had shown that the mixed culture accelerated proteolysis significantly. Thus, two-dimensional electrophoresis confirmed the hydrolysis of actin, myosin light chain 1/3, myosin light chain 2, and myosin heavy chain. 33 peptides from troponin T, myosin light chain 2, and, in particular, from actin were also identified by means of LC-MS/MS. In the matter of actin primary structure, three of its regions were very sensitive to degradation. Although the inoculation of the mixed starter cultures accelerated proteolysis, peptides and amino-acids contents were reduced while L. curvatus CRL705 alone increased the abovementioned contents.
In 2018, De Almeida et al. [3] evaluated the ability of four species of lactic acid microorganisms to form low molecular weight peptides in a sausage model with a low sodium content. As a whole, 86 low molecular weight peptides obtained mainly from myofibrillar proteins were identified (56 of them – from actin). The inoculation of L. curvatus CRL705 and Enterococcus mundtii CRL35 led to the formation of 56 and 43 low molecular weight peptides, respectively. Larger quantity of peptides per unit time was released when using Lactobacillus plantarum and L. sakei in comparison to other models. Each strain in the models allowed creating the unique profile of small peptides and amino acids which might be used as a biochemical characteristic to differentiate certain fermented foods.
In the research conducted by Basso et al. [4], Lactobacillus sakei (DSM 6333), Lactobacillus plantarum (B21), and L. farciminis (DSM 20184) have demonstrated proteolytic activity to meat sarcoplasmic proteins. Thus, the 94kDa-band disappeared in samples with L. farciminis and L. plantarum and narrowed significantly in samples with L. sakei. Bands with a molecular mass from 94 kDa to 38 kDa decreased in all samples. According to capillary electrophoresis analysis, no signals corresponding to the retention time of 8.64 and 8.66 minutes were observed in all the samples. Moreover, MALDI-TOF MS method identified 94-kDa and 38-kDa bands as an isoform of muscle glycogen phosphorylase and glyceraldehyde-3-phosphate dehydrogenase.
In 2001, Italian researchers [5] obtained controversial results on the proteolytic activity of 27 Staphylococcus xylosus strains to sarcoplasmic and myofibrillar proteins. Neither electrophoretic nor spectrometric analysis confirmed the proteolytic activity of some strains. SDS-polyacrylamide gel electrophoresis revealed changes in pork protein profile for only 12 out of 24 strains which were able to hydrolyze muscle protein extracts on agar.
Montowska et. al. [6] has proved a favorable effect of trypsin along with super-high frequency treatment on rapid and efficient recovery of peptide markers in meat products. Meat was subjected to heating – considerable protein aggregation was observed – and then analyzed by means of tandem mass spectrometry to detect specific peptide markers. It was established that treatment of heat-treated product with trypsin for one hour contributes to the structure recovery of desirable peptide markers which could be easily identified using the known methods.
Ageing effects meat taste, tenderness, water-binding capacity (WBC), colour, and juiciness. The monitoring of biochemical processes occurring during meat ageing allows meat products quality biomarkers to be revealed [7]. Change in the proteome of muscle exudate from genetically similar pigs with the same meat characteristics within a regular ageing period, namely, 7 days, was studied [1]. It has been found that some quality characteristics of meat such as meat tenderness, cooking losses as well as the colour index CIE b change considerably due to autolysis, notably, at the end of the ageing process. These data illustrated structural changes in the pork meat during the ageing process, which affected proteomic profiles. Three main groups of proteins which changed significantly during the ageing were detected: stress proteins, metabolic enzymes, and structural proteins. Undoubtedly, proteolysis plays a key role in protein fragmentation (for instance, enolase and titin). It has been also observed that the number of stress proteins increase. To monitor these changes, myofibrillar or sarcoplasmic proteomic fractions are usually used. However, using the muscle exudate as a more available substrate allowed previous research to be expanded. For example, it has been discovered that vinculin and peroxiredoxin-6 correlate to the WBC and the tenderness of meat, respectively. Such protein biomarkers are useful for monitoring the fresh meat quality and forecasting the autolysis course.
Oxygen, contained in meat tissues, also takes part in the formation of meat raw materials texture, namely, tenderness and juiciness: a high oxygen level contributes to protein intermolecular cross-linking and aggregates formation [9, 10].
The results of the research conducted by Moszkowska et. al. [11] have shown the formation of protein aggregates in the muscle tissue packaged in an oxygen-containing atmosphere. These aggregates, which consist of myosin heavy chains, are the result of cross-linking proteins and are able to affect the meat tenderness adversely. Aggregate formation depends not only on a type of packaging but also on a method of packaging. Thus, larger aggregates were observed in biceps femoris muscles in comparison to longissimus lumborum muscles and in meat packaged in the modified gas atmosphere (MGA) rather than in vacuum. It may be explained by the metabolism of muscles analyzed due to differences in postmortem glycolysis extent, the characteristics of muscle fibers and their glycolytic and oxidative properties. Both SDS-gel electrophoresis and diagonal-PAGE, as well as immunoblotting, demonstrated the oxidative cross-linking of myosin heavy chains in meat packaged in MGA with high oxygen content. Moreover, aggregates of myosin heavy chains (MHC) were also detected in other packages containing high oxygen concentration (MGA and vacuum + MGA); apparently, protein oxidation occurred which led to myosin binding due to its polymerization [12]. Such an aggregation of MHC, occurred under severe temperature conditions (190°C), and took place in both raw meat and finished meat products [6].
The aggregate formation can be related not only to oxidation but to proteolysis as well. In this work, we identified a number of protein aggregates of different origin formed as a consequence of proteolytic and oxidative processes.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
In this research, we studied m. longissimus dorsi samples from Bos Taurus before and after treatment with proteolytic enzyme of animal and plant origin and a number of starters. Whole muscle of 500 g was injected with proteases solutions in the amount of 5 ml per 50 g raw meat and kept at 30°C for 40 min for trypsin and pepsin, and for 30 min for papain and bromeline treatment. In case of starter cultures, a sample was kept in vacuum at 11°C for 9 days.
We used 1.5% pig pepsin and bovine trypsin solutions (Himedia, India, 10, 000 NFU/mg and 2, 000 U/g, respectively) and 0.5% papain and bromeline solutions (Sigma, USA, 1.1 U/mg solid and 2, 370 U/g prot., respectively). The starter suspensions concentration was determined as 109 CFU/ml using McFarland standard. The following starters were used: Pediococcus pentosaceus 31 (VKPM-8901), Pediococcus acidilactici 38 (VKPM-8902), Lactobacillus sakei 105 (VKPM-8905), and Lactobacillus curvatus 2 (VKPM-8906).
Another set of experiments was carried out using uncooked smoked sausage samples. The control sample did not include starters. The experimental sample no. 1 had starters from the Microorganisms Collection of Moscow State University of Food Production. The experimental sample no. 2 was prepared using starters from Bactoferm SM 194 Chr. Hansen, Denmark, such as Pediococcus pentosaceus, Staphylococcus carnosus, Staphylococcus xylosus, Lactobacillus sake, and Debaryomyce shansenii. Uncooked smoked sausage for the experiment was manufactured by a halal meat production factory in the Moscow region. It included such ingredients as beef of best quality, beef and horse meat of good quality, soy granules, beef external fat, vegetable fiber with a coloring agent, nitrite salt, and spices.
In the third set of experiments, skeletal muscles of Bos Taurus (m. longissimus dorsi) were separated from a carcass in 30 min after slaughter and vacuum-packaged into individual packages. The samples were kept at ± 2°C for 4 and 7 days until analyzed.
Protein fractionation, colouring, computer densitometry, and mass-spectrometric identification were carried out as described in previous research [13, 14]. Protein identification was financially supported by the Ministry of Education and Science of the Russian Federation and performed in Shared Knowledge Center ‘Industrial biotechniques’ (RFMEFI62114X0002).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
The enzyme preparations listed are widely used in the meat industry. For instance, the American Food and Drug Administration of the Department of Health Protection and Human Services generally recognized five exogenous enzymes as safe (GRAS) to use in the meat industry. These are papain, bromeline, ficin, and Aspergillus oryzae and Bacillus subtilis proteases [15]. Therefore, a potential formation of complexes limiting the exoprotease usage due to their reduced effectiveness in meat is of great interest.
The silver nitrate staining of a bovine skeletal muscle treated with trypsin and pepsin revealed a protein track with molecular mass of 15–100 kDa in the alkaline zone of pH gradient (Fig. 1). The track was not detectable in staining with CBB R-250. The former staining technique is 100 times more sensitive than the latter one, which means a small quantity of the protein involved. The analysis of gel fragments from upper and lower zones of this track revealed isoform 2 of protein 1 containing 4.5 LIM domains, FHLI gene. The monomeric form of the protein corresponded completely to its physical and chemical parameters (molecular mass and pH) and contained more than 90% of protein substances in the track.

The amino acid sequence of the protein does not exceed 32 kDa in mass, whereas that in the track was considerably wider. It implies that the proteolytic treatment causes aggregates formation that gives an unusual electrophoretic track. The analysis of vacuum-packaged skeleton muscles of Bos Taurus also detected an identical track of the protein by day 4 of storage that decreased by day 7 as a consequence of the autolytic process. After unpacking, the trace formation process tended to accelerate during 3 days, which confirms that both proteolytic enzymes and free oxygen take part in this process.
The track formation can be explained by the fact that animal proteases act in injection sites, and the stage of aggregates formation is impossible to record. However, one of the proteolysis stages is the removing of similar molecules. This implies that a track with reduced mass is formed which can be detected only at low protease concentrations, i.e. at some distance from injected sites. Plant proteases do not have such an effect, probably, because their pH optimums are more adapted to raw meat conditions.
Thus, bromeline resulted in the formation of five high molecular fractions of myosin heavy chains fragments with molecular weights of 170, 150, 60, and 44 kDa, respectively. Consequently, polypeptides with a high molecular weight, a few short peptides, and a small number of free amino acids are formed as a result of bromeline action. Bromeline has a low actomyosin affinity. The data resulted coincide with those of Istrati et. al. [8].
Another case of protein aggregates formation was detected when using Pediococcus pentosaceus 31 VKPM-8901as a starter. Fig. 2 demonstrates weight high molecular protein aggregates (160–200 kDa) that were identified as myoglobin and troponin (Table 1). Their subunits molecular weights did not exceed 21.5 kDa, and their monomers were represented in large quantities on the two-dimensional electrophoregram. In addition, fractions with much higher weight (up to 3% of monomers) were also recorded.
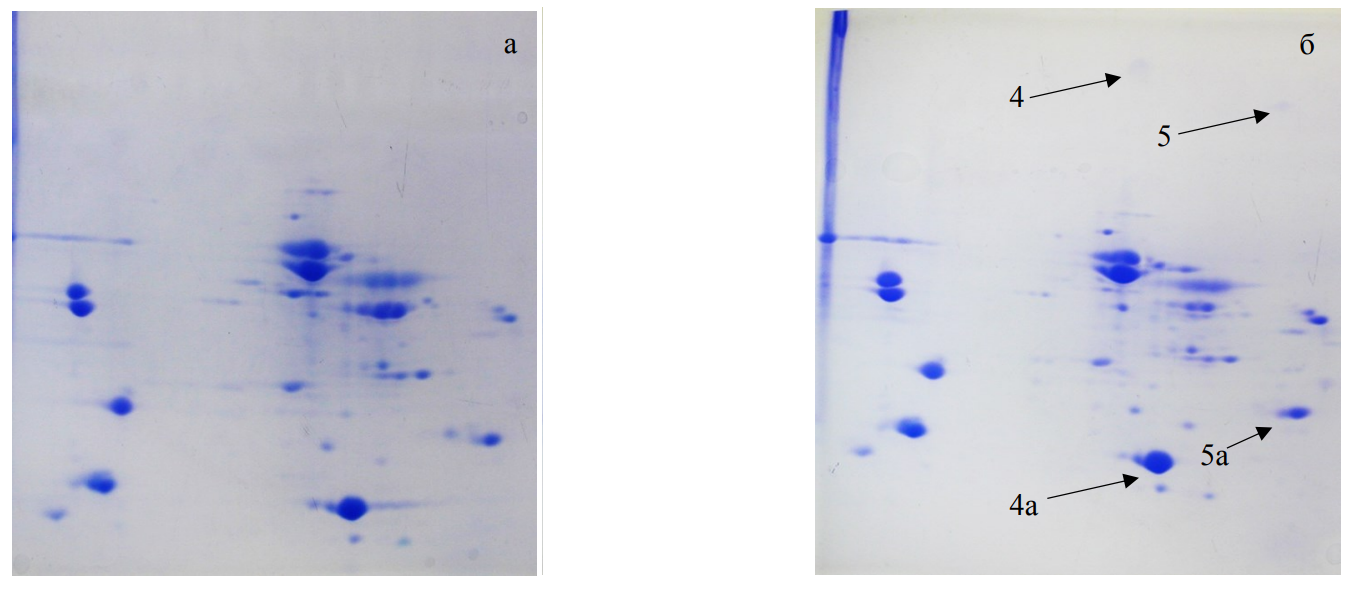
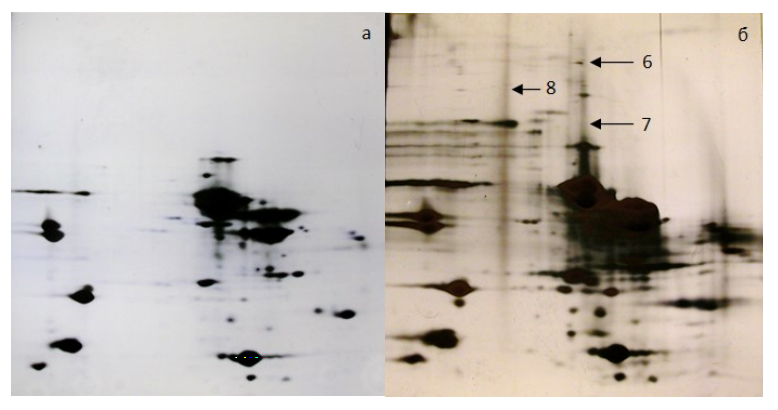
In vacuum-packaged skeletal muscle from Bos Taurus, aggregates of other proteins were also revealed (Fig. 3). Aggregates of mixed fragments, such as muscle creatine phosphokinase and glutathione-S-transferase, actin and perilipin, and type II keratin, appeared. A cause of their formation might be in a low oxygen concentration and naturally occurring autolysis. These factors slow down the aggregates formation process through the removal of structurally similar molecules complex from submolecular structures, which can be seen on the electrophoregrams.
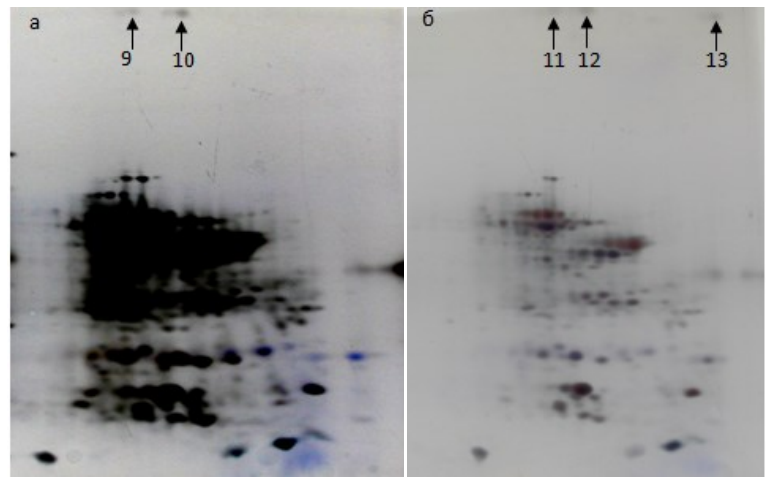
Aggregation was also observed in uncooked smoked sausage samples with starters from the MSUFP collection (Fig. 4). Two of the samples contained soy protein aggregates (soy was used in the sausage formulation). The control sample (without starters) included two aggregates of glycin G1 fragments. The sample with starters had glycin G1 fragments and a glycinin A3B4 aggregate, and the third sample did not contain aggregates. These results demonstrate that the presence of oxygen and the use of several types of proteases make aggregates formation with plant proteins possible.
ВЫВОДЫ
According to the data obtained, only certain proteases provoked protein aggregation, and the presence of free oxygen accelerated the process. The proteases affected several types of raw meat proteins and also plant proteins in meat products (in particular, soy ones). Additionally, some proteins were also involved but one-dimensional electrophoresis was not able to detect them because of small quantities. Those proteins had alkali pI, therefore the use of immobilized pH gradients was useless to detect them as opposed to ampholines non-equilibrium pH gradient electrophoresis. The results obtained, along with those of previous research [12, 14], allowed us to assume that cross linkages formation and the aggregation of muscle proteins, primarily myosin, tended to reduce meat water-binding capacity, juiciness, and tenderness and affected meat flavor adversely. The negative impact of protein polymerization on the calpain activity also might retard the tenderization process in early proteolysis in the presence of free oxygen [12].
Apparently, those proteases which led to protein aggregate formation were less effective for meat tenderization and formation of consumer characteristics of finished meat products.
КОНФЛИКТ ИНТЕРЕСОВ
The authors declare no conflict of interest.
ФИНАНСИРОВАНИЕ
The study was financially supported by the Russian Science Foundation (Project No. 16-16-10073).
СПИСОК ЛИТЕРАТУРЫ
- Di Luca A., Elia G., Mullen A.M., and Hamill R.M. Monitoring post mortem changes in porcine muscle through 2-D DIGE proteome analysis of Longissimus muscle exudate. Proteome Science, 2013, vol. 11, no. 1. DOI: https://doi. org/10.1186/1477-5956-11-9.
- López C.M., Sentandreu M.A., Vignolo G.M., and Fadda S.G. Proteomic and peptidomic insights on myofibrillar protein hydrolysis in a sausage model during fermentation with autochthonous starter cultures. Food Research Inter- national, 2015, vol. 78, pp. 41–49. DOI: https://doi.org/10.1016/j.foodres.2015.11.009.
- De Almeida M.A., Saldaña E., da Silva Pinto J.S., et al. A peptidomic approach of meat protein degradation in a low-sodium fermented sausage model using autochthonous starter cultures. Food Research International, 2018, vol. 109, pp. 368–379. DOI: https://doi.org/10.1016/j.foodres.2018.04.042.
- Basso A.L., Picariello G., Coppola R., et al. Proteolytic Activity of Lactobacillus Sakei, Lactobacillus Farciminis and Lactobacillus Plantarum on Sarcoplasmic Proteins of Pork Lean. Journal of Food Biochemistry, 2004, vol. 28, no. 3, pp. 195–212. DOI: https://doi.org/10.1111/j.1745-4514.2004.tb00066.x.
- Mauriello G., Casaburi A. and Villani F. Proteolytic activity of Staphylococcus xylosus strains on pork myofibrillar and sarcoplasmic proteins and use of selected strains in the production of “Naples type” salami. Journal of Applied Microbiology, 2002, vol. 92, no. 3, pp. 482–490. DOI: https://doi.org.10.1046/j.1365-2672.2002.01551.x
- Montowska M. and Pospiech E. Processed meat protein and heat-stable peptide marker identification using mi- crowave-assisted tryptic digestion. Food Technology and Biotechnology, 2016, vol. 54, no. 4, pp. 482–488. DOI: https://doi.org/10.17113/ftb.54.04.16.4540.
- Zamaratskaia G. and Li S. Proteomics in meat science – current status and future. Theory and Practice of Meat Pro- cessing, 2017, vol. 2, no. 1, pp. 18–26. (In Russ.). DOI: https://doi.org/10.21323/2414-438X-2017-2-1-18-26.
- Istrati D., Vizireanu C., and Dinica R. Influence of post-mortem treatment with proteolytic enzymes on tenderness ofbeef muscle. Journal of Agroalimentary Processes and Technologies, 2012, vol. 18, no 1, pp. 70–77.
- Morzel M., Gatellier P., Sayd T., Renerre M., and Laville E. Chemical oxidation decreases proteolytic susceptibility of skeletal muscle myofibrillar protein. Meat Science, 2006, vol. 73, no. 3, pp. 536–543. DOI: https://doi.org/10.1016/j. meatsci.2006.02.005.
- Lund M.N., Lametsch R., Hviid M.S., Jensen O.N., and Skibsted L.H. High-oxygen packaging atmosphere influences protein oxidation and tenderness of porcine longissimus dorsi during chill storage. Meat Science, 2007, vol. 77, no. 3, pp. 295–303. DOI: https://doi.org/10.1016/j.meatsci.2007.03.016.
- Moczkowska M., Półtorak A., Montowska M., Pospiech E., and Wierzbicka A. The effect of the packaging system and storage time on myofibrillar protein degradation and oxidation process in relation to beef tenderness. Meat Science, 2017, vol. 130, pp. 7–15. DOI: https://doi.org/10.1016/j.meatsci.2017.03.008.
- Kim Y.H., Huff-Lonergan E., Sebranek J.G., and Lonergan S.M. High-oxygen modified atmosphere packa- ging system induces lipid and myoglobin oxidation and protein polymerization. Meat Science, 2010, vol. 85, no. 4, pp. 759–767. DOI: https://doi.org/10.1016/j.meatsci.2010.04.001.
- Kovalyov L.I., Shishkin S.S., Kovalyov M.A., et al. Proteomic research proteins in a sample of pork meat products.All about the meat, 2013, no. 3, pp. 32–34. (In Russ.).
- Zvereva E.A., Kovalev L.I., Ivanov A.I., et al. Enzyme immunoassay and proteomic characterization of troponin I as a marker of mammalian muscle compounds in raw meat and some meat products. Meat Science, 2015, vol. 105, pp. 46–52. DOI: https://doi.org/10.1016/j.meatsci.2015.03.001.
- Generally Recognized as Safe (GRAS). Available at: https://www.fda.gov/Food/IngredientsPackagingLabeling/ GRAS/ucm2006850.htm. (accessed August 2018).
- Chen L., Zhou G.-H., and Zhang W.-G. Effects of high oxygen packaging on tenderness and water holding ca- pacity of pork through protein oxidation. Food and Bioprocess Technology, 2015, vol. 8, no. 8, pp. 2287–2297. DOI: https://doi.org/10.1007/s11947-015-1566-0.
