

Abstract
Introduction. Recent studies have shown the benefits of phytolytic enzymes to prepare grain wort in ethanol production. However, there is a lack of data on the effect of phytases and their amount on the conversion of grain polymers, the ionic composition of wort and mash, and the efficiency of yeast generation and ethanol fermentation.Study objects and methods. Wheat and corn wort samples were treated with a complex of hydrolases, including phytases. Capillary electrophoresis determined the ionic composition of wort and mash. Gas chromatography measured the content of volatile metabolites.
Results and discussion. The key enzymes were phytases and proteases. They improved the conversion of grain polymers and stimulated the growth and metabolism of yeast cells. Their synergism enriched the wort with assimilable nitrogen, phosphorus, and other valuable minerals. In addition, it intensified the growth of the Saccharomyces cerevisiae yeast, increased the rate of carbohydrate consumption, and reduced the formation of side metabolites 1.7–1.9 times, mainly due to higher and aromatic alcohols. The concentration of phosphates remained practically unchanged during the fermentation of grain wort treated with phytases. However, by the end of fermentation, it was 2.4–5.1 times higher than in the mash samples without phytolytic treatment. Finally, we identified a complex of enzymes and optimal amounts of phytases that have a stimulating effect on ethanol fermentation.
Conclusion. Phytases, whether used individually or together with proteases, enriched grain wort with soluble macro- and microelements, improved yeast metabolism, directed ethanol synthesis, and decreased the formation of fermentation by-products.
Keywords
Wort, phytase, protease, mash, yeast, metabolism, ethanol fermentationINTRODUCTION
Recent studies have proven the effectiveness of mechanical and enzymatic treatment of grains (at temperatures under 100°C) with complex enzyme preparations in ethanol production [1–4]. This “soft” technology for grain wort preparation can significantly reduce heat and power consumption and increase profitability. It is based on controlled biocatalytic conversion of grain polymers (starch, xylans, β-glucans, and proteins) with the formation of easily digestible carbohydrates and nitrogenous substances that yeast cells need for normal metabolism [2, 4–7].
However, the studies hardly took into account the presence of phytic acid and its salts (phytates) in grains that contain up to 80% of phosphorus in a bound state [8–10]. The bioavailability of phosphorus can be increased by using phytolytic enzymes. Grains contain enzymes that catalyze the destruction of intracellular polymers. Normally in a latent state, these enzymes are activated during germination. Studies show that the amount of phytolytic enzymes in grain is insufficient for the complete release of phosphorus [11, 12]. Therefore, researchers have recently focused on obtaining enzyme preparations – sources of phytases – based on microorganisms. Using genetic engineering, they have developed highly productive recombinant strains of fungi, yeasts, and bacteria that synthesize phytolytic enzymes [13–15].
Phytolytic enzymes of microbial origin have been widely studied, namely their use in fodder production for the release of phytate phosphorus. The biocatalytic conversion of plant phytates has been shown to improve the digestibility of nutrients in fodder and stimulate the growth of farm animals and poultry [16–18]. Much attention has been paid to the use of phytases in the food industry to increase the digestibility and bioavailability of food components [19–21]. However, there is a lack of data concerning the catalytic action of microbial phytases on grain polymers in fermentation.
Previous studies have shown a positive effect of phytases on the growth and development of yeast cells cultivated on grain media, including beer production from sorghum [22–24]. Some authors who studied the effect of phytolytic enzymes reported better rheological properties of rye and triticale wort, as well as improved fermentation [6, 25, 26].
However, there is a lack of research into their effect on the biochemical parameters of grain wort and mash, especially those from wheat and corn [2, 23]. What needs studying is changes in the ionic composition of grain wort depending on the substrate specificity of enzyme systems used for its preparation, as well as the assimilation of the released phytate phosphorus by the Saccharomyces cerevisiae yeast during ethanol fermentation [1–3, 22–25]. Hardly studied is the effect of phytases on the hydrolytic capacity of enzymes with substrate specificity in relation to starch and proteins, on the conversion of these polymers in grain wort preparation, and on the efficiency of yeast generation and ethanol fermentation.
Thus, literature analysis revealed a lack of studies into the potential of phytic substances with bound phosphorus for the conversion of grain to ethanol. A number of papers reported that phytates – strong chelating agents – bind not only phosphorus, but also metal cations (calcium, magnesium, manganese, zinc, iron, etc.) [19, 21, 27–29]. Low bioavailability of macroand microelements in grains can have a negative effect on the supply of yeast with minerals and on the catalytic activity of some metal-dependent enzymes.
The S. cerevisiae yeast is a key factor in the efficiency of ethanol fermentation. The yield and quality of ethanol, as well as the process duration, largely depend on the fermentation activity and yeast productivity. The metabolism of S. cerevisiae is significantly affected not only by the strains’ genetic characteristics, but also by the conditions of their cultivation (nutrient medium with easily assimilated nutrition) [2, 30–32].
Metabolism involves all enzymatic reactions that occur in the cell to regulate the composition and synthesis of target and secondary metabolites. Therefore, there is a need for research to select effective enzyme systems that contribute to deep destruction of grain polymers. It is especially true of phytolytic enzymes. Recent studies have revealed that phytic acid forms stable complexes with carbohydrates and proteins [33–35]. Apparently, this can reduce the hydrolytic action of carbohydrases and proteases on carbohydrate and peptide polymers. Phosphate groups of phytic acid bind to basic amino acids (arginine, histidine, and lysine) and form strong protein-phytate complexes.
However, the studies of phytate-carbohydrate complexes have ambiguous results. They show that the interaction occurs either through direct binding to starch or indirectly, through starch-associated proteins [34–37]. Therefore, to ensure a steadily high yield of ethanol, it is important to select optimal parameters for preparing grain media to produce high-quality wort [1–3].
In connection with the above, we can assume that the biocatalytic destruction of phytic substances will contribute to the release of phosphorus and other valuable microelements. It will also stimulate the conversion of carbohydrate and protein polymers of grain through the use of hydrolases with substrate specificity in relation to the main polymers of grain.
We aimed to study the effect of hydrolytic enzymes with proteases and phytases on the efficiency of yeast generation and metabolism during the fermentation of wheat and corn wort.
STUDY OBJECTS AND METHODS
Our study objects included wheat and corn wort
prepared with enzyme preparations that served as
sources of hydrolases with different substrate specificity
and action. They were used for:
‒ dextrinization and saccharification of starch α-amylase
(EC 3.2.1.1.) and glucoamylase (EC 3.2.1.3.);
‒ destruction of xylanase non-starch polysaccharides
(EC 3.2.1.8, 3.2.1.32, 3.2.1.37, 3.2.1.72);
‒ hydrolysis of protein substances of the protease
complex (EC 3.4.11-3.4.15, EC 3.4.21-3.4.24); and
‒ hydrolysis of phytase substances (EC 3.1.3.8).
Yeast, organic acids, and dietary supplements were obtained from the Biotechnology Department. Ultraconcentrates of culture liquids were used as enzyme preparations to produce thermostable α-amylase (Bacillus licheniformis sp., Amilolikheterm), glucoamylase and xylanase (VKM F-4278D, a recombinant strain of Aspergillus awamori, Glucavamorin-Xyl), a protease complex (VKPM F-931, a mutant strain of Aspergillus oryzae, Protoorizin), and phytase (Phytaflow, Novozymes, Denmark) [38–40].
Enzyme activity was determined by the existing methods. A unit of amylolytic activity (AA) was defined as an amount of enzyme that catalyzes the hydrolysis of 1 g of soluble starch to dextrins of various molecular weights under standard conditions (30°C, pH 6.0, 10 min). A unit of glucoamylase activity (GA) was an amount of enzyme that is capable of catalyzing starch hydrolysis at 30°C (pH 4.7) and releasing 1 μmol of glucose per minute. A unit of xylanase activity (XA) was an amount of enzyme that acts on xylan from birch wood and releases 1 μmol of reducing sugars (in glucose equivalent) per minute under standard conditions (50°C, pH 5.0). A unit of total proteolytic activity (PA) was an amount of enzyme that brings hemoglobin into a TCA non-precipitated state equivalent to 1 μmol of tyrosine per minute (30°C, pH 5.3). A unit of phytase activity (PhA) was an amount of enzyme that catalyzes the hydrolysis of sodium phytate to produce 1 μmol of inorganic phosphate per minute under standard conditions (37°С, pH 5.5, 15 min).
Grain wort was prepared by the enzymatichydrolytic processing of grain (40–90°C, hydromodule 1:3). For this, 50 g of grain flour and 150 cm3 of water were mixed in 750 cm3 Erlenmeyer flasks and regularly stirred in a PE-4300 water bath (Ekros, Russia).
The water-grain mixture was prepared for 2 h (40–90°C) using a thermostable α-amylase (0.6 AA units/g starch) to liquefy starch. When the mixture was cooled to 60°C (56–60°C, 1 h), EPs with glucoamylase (9.0 GA units/g starch) and xylanase (0.4 XA units/g grain) activity were used to saccharificate starch and hydrolyze non-starch polysaccharides. They served as a control. The experimental samples contained EPs with phytase (1.0–3.0 PhA units/g grain) and proteolytic activity (0.1 PA units/ g grain), in addition to those with amylolytic, glucoamylase, and xylanase activity.
Wort was fermented using a selected race of the Saccharomyces cerevisiae 985-T yeast with thermotolerant and osmophilic properties (34–35°C, 68 h) [41].
Methods. The Instructions for the Technochemical Control of Alcohol Production were followed to determine the contents of starch, protein, and nonstarch polymers of grain, the concentrations of yeast cells and reducing carbohydrates in the grain mash, as well as ethanol concentration and yield [42]. The concentration of amine nitrogen in the grain wort was measured by iodometric titration (Pharmaceutical Regulations 1.2.3.0022.15). The ionic composition of wheat and corn wort, as well as mash, was determined using a PrinCE-560 capillary electrophoresis system (Netherlands) equipped with a conductometric detector. The composition and content of volatile metabolites synthesized by yeast were measured using an HP Agilent 6850 gas chromatograph (USA).
Data obtained in triplicate were statistically processed in Microsoft Excel using the Student’s coefficient with a 0.95 confidence interval.
RESULTS AND DISCUSSION
Ground wheat and corn were used as a substrate to prepare wort. Table 1 compares the compositions of the main polymers contained in the grains. According to their caryopsis composition, wheat and corn, like all grains, are classified as multicomponent starchy plant raw materials, in which starch is the main polymer that determines ethanol yield [10, 25, 42].
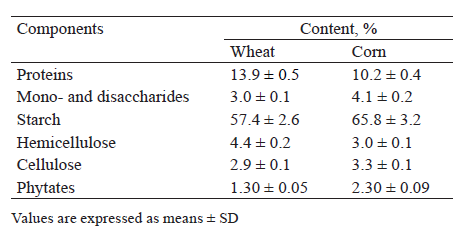
Corn had a higher content of starch (65.8%), while wheat was richer in protein (13.9%). In addition, the grains under study contained non-starch polysaccharides (hemicelluloses, cellulose), protein substances, and phytates (Table 1). This multicomponent composition of grain polymers determined the selection of enzyme preparations with a given substrate specificity to prepare grain wort (Table 2). Amilolicheterm (thermostable α-amylase, 330 AA units/g) was used to liquefy and dextrinize starch. Glucavamorin-Xyl (glucoamylase, 7700 GA units/g; xylanase, 350 XA units/g) was used to saccharify starch and hydrolyze non-starch polysaccharides. Protoorizin (protease, 580 PA units/g) was used for protein proteolysis and Phytaflow (30 000 PhA units/g) was selected to convert phytic substances.
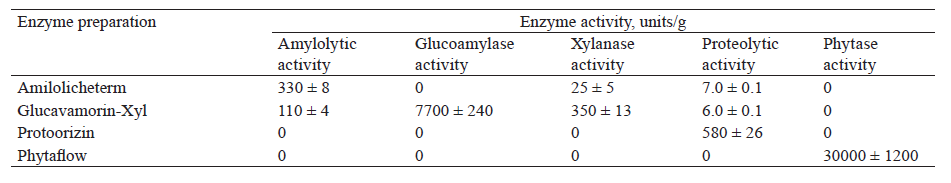
Protoorizin contained at least five proteolytic enzymes that differ in action [39]. In previous studies, the authors used neutral protease or Glucavamorin-Xyl, which contained proteinases to catalyze the hydrolysis of proteins to peptides with different molecular weights [24, 25]. In contrast to them, Protoorizin contains a complex of proteinases and peptidases that hydrolyze proteins to low-molecular-weight peptides and free amino acids [40, 43].
Enzyme preparations were used at the stage of grain wort preparation in the amounts specified above (see “Study objects and methods”). The control wort contained Amilolicheterm and Glucavamorin-Xyl preparations, the sources of carbohydrases that catalyze the hydrolysis of polysaccharides. Their proteolytic enzymes had practically no effect on the degree of protein conversion due to the low level of their activity. The concentration of proteases in the control was under 0.02 PA units/g grain, which was 5 times lower than in the experimental samples. Therefore, Protoorizin (0.1 PA unit/g grain) and Phytaflow (1.0–3.0 PhA units/g grain) were additionally added to the experimental samples to ensure an efficient conversion of grain polymers and activate yeast generation and ethanol fermentation.
We found that the ionic composition of grain wort changed depending on the type of grain and the substrate specificity of the enzymes used to hydrolyze grain polymers. In the wheat wort treated with a complex of amylolytic and xylanase enzymes (AA + GA + XA), the phosphate content was 2.7 times higher than in the corn wort (Fig. 1). The concentrations of other cations, such as potassium, calcium, sodium, and magnesium, were also different. The wheat wort was richer in potassium, while the corn wort had a higher content of calcium and magnesium (Table 3).
Phytolytic enzymes had a more significant effect on the release of phytate phosphorus. Increasing their amount to 1.5–2.0 PhA units/g grain led to higher concentrations of released phosphates due to the catalytic destruction of phytic substances. We found that the content of phosphorus ions increased 1.6 times in the wheat wort and 3.8 times in the corn wort. A further increase in the phytase concentration to 3.0 PhA units/g grain had practically no effect on the content of soluble phosphates in the wort (Fig. 1).

The catalytic synergism of phytases and proteases stimulated the release of phosphorus ions. Their content increased 1.8 times in the wheat wort and 4.9 times in the corn wort (Table 3).
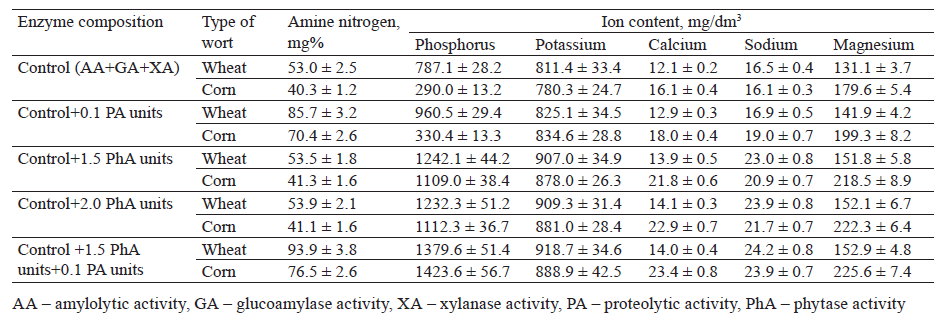
In addition to the release of phosphorus ions, phytases increased the concentration of cations in the wheat and corn wort samples: potassium by 12 and 13%, calcium by 15 and 35%, sodium by 39 and 30%, and magnesium by 16 and 22%, respectively (Table 3).
We found that the content of cations depended on the substrate specificity of the enzymes involved in the bioconversion of grain polymers. The combined action of proteolytic and phytolytic enzymes contributed not only to the accumulation of phosphates and minerals, but also to a significant increase in amine nitrogen. Its concentration was 1.8 times as high in the wheat wort and 1.9 times as high in the corn wort, compared to the control.
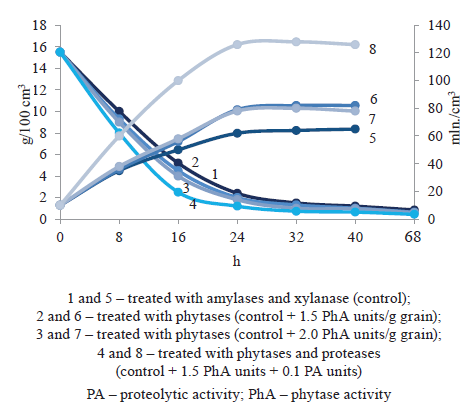
Grain wort enriched with phosphates and minerals was used as a nutrient medium to cultivate the ethanol yeast Saccharomyces cerevisiae (race 985-T). We studied the process of yeast generation on grain wort prepared with hydrolytic enzymes differing in substrate specificity and found that the presence of phytase in the enzyme complex had a positive effect on the growth and reproduction of yeast cells (Fig. 2).
As we know, mineral and nitrogenous substances are essential for the biochemical reactions of yeast cells [24, 25, 43]. Magnesium and calcium ions activate the catalytic ability of almost all intracellular metalloenzymes, including phosphofructokinase, which is involved in glucose metabolism. Potassium, sodium, and calcium ions have a regulatory effect on the metabolism of yeast cells. Potassium also plays an essential role in oxidative phosphorylation and glycolysis processes [43]. It activates yeast aldolase and, together with magnesium ions, catalyzes pyruvate carboxylase. Like nitrogen, potassium can also affect yeast lipid metabolism.
We analyzed the processes of yeast generation and carbohydrate consumption and found that the presence of phosphorus and other minerals in the medium intensified the growth of S. cerevisiae yeast. In the lag phase (first 18–24 h of growth), the concentration of yeast cells increased 1.2–1.3 times, alongside a rising rate of carbohydrate consumption. A 1.3-fold increase in phytolytic enzymes (up to 2.0 PhA units/g grain) during grain wort preparation had no significant effect on the yeast growth and the assimilation of reducing carbohydrates (Fig. 2).
The catalytic synergism of phytase and protease significantly enriched the wheat wort with mineral and nitrogenous nutrition and resulted in the most active growth of yeast cells (2-fold) and a more intensive consumption of carbohydrates (Fig. 2, curves 4 and 8). A similar pattern was observed with the corn wort.
Thus, our results confirmed that the nutrient medium has a significant effect on yeast generation and physiological activity, particularly the presence of soluble macro- and microelements in addition to easily digestible carbohydrates and nitrogenous substances.
At the next stage, we analyzed changes in the concentration of phosphates in the mash against the amount of phytolytic enzymes used in the preparation of wheat and corn wort (Figs. 3a and 3b). We found that the content of phosphorus ions significantly decreased during the fermentation of the control wort, which was not treated with phytases. In the logarithmic phase of yeast cell growth (on day 1), it declined 1.9 times in the wheat wort and 2.3 times in the corn wort. The wort samples treated with protease in addition to carbohydrases showed the same trend – a sharp decrease in phosphates after 24 h of fermentation, followed by a slight rise.
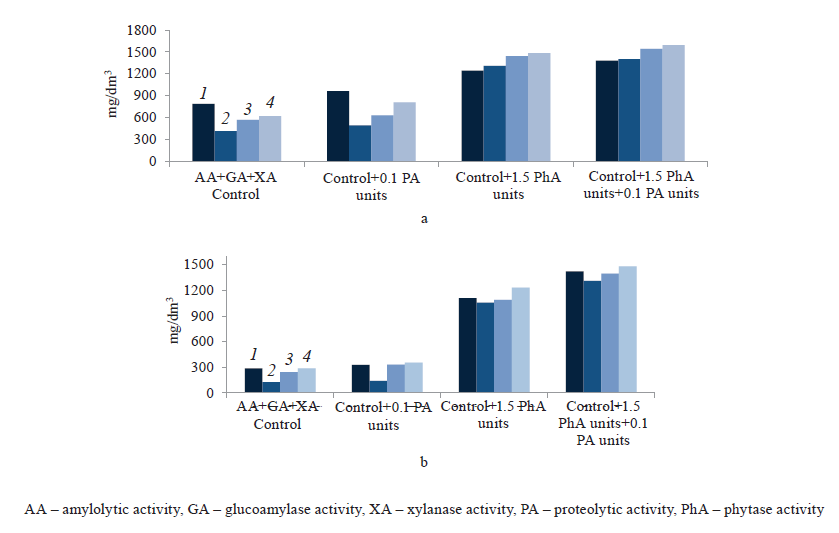
In the experimental samples treated with phytolytic enzymes, the concentration of phosphates hardly changed during the fermentation of grain wort. This might be due to the ongoing biocatalytic hydrolysis of phytic substances and an extra release of phosphorus. By the end of fermentation, the content of phosphates slightly increased, which might be associated with autolytic processes in the cell (Fig. 3).
We found an increase of 2.4–2.6 times and 4.3–5.1 times in the residual content of phosphates in the wheat mash and the corn mash, respectively. This indicated that phytolytic enzymes not only enriched the grain wort with assimilable phosphorus and other valuable minerals, but also improved the value of grain stillage, a waste product of ethanol production that is used in the diet of farm animals.
Apart from the main fermentation products (ethanol and carbon dioxide), yeast cells synthesize accompanying metabolites: secondary (organic acids, aldehydes, and esters) and by-products (higher alcohols) [43].
We studied the metabolism of S. cerevisiae 985-T yeast during its cultivation on wheat and corn wort treated with phytolytic enzymes and found a decrease of 18–21 and 20–23%, respectively, in total metabolite formation that accompanies ethanol synthesis (Fig. 4). These results indirectly confirmed the improvement of phosphorus metabolism in the cell.
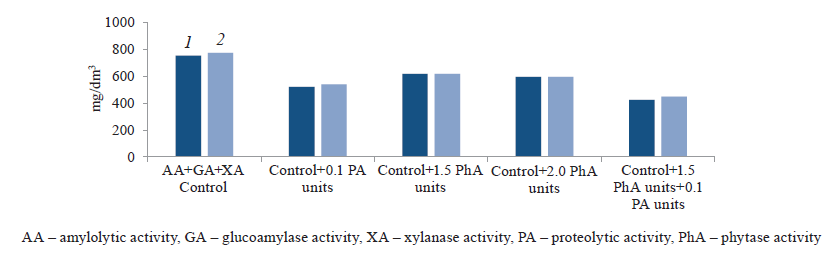
The use of proteases reduced the synthesis of side and secondary metabolites by more than 30%. The greatest effect was caused by the combined catalytic action of phytases and proteases during yeast generation on the media enriched with assimilable phosphates and amine nitrogen. In particular, the total content of accompanying volatiles in the wheat and corn mash decreased by 44 and 42%, respectively, compared to the control.
Higher alcohols (mostly isoamylol, isobutanol, and 1-propanol) dominated among side and secondary metabolites synthesized by S. cerevisiae 985-T during the wort fermentation. In addition, the mash contained aromatic alcohols (β-phenylethyl, p-hydroxyphenylethyl) and a small amount of secondary by-products, primarily aldehydes and esters (Fig. 5).

Our study showed that treating the nutrient media with phytolytic and proteolytic enzymes decreased the synthesis of higher and aromatic alcohols (1.9 and 1.4 times, respectively) (Fig. 5). Exposure to phytases reduced the amount of higher alcohols and proteases (1.3–1.4 and 1.5–1.6 times, respectively). In addition, we observed a slight decrease in aldehydes and esters.
Thus, our study showed that providing a yeast cell with a balanced nitrogenous and mineral nutrition created conditions for synthesizing ethanol with a reduced amount of fermentation by-products. By regulating yeast metabolism we can improve the quality and sensory properties of the target product.
According to our results, the biochemical changes in the grain wort affected the yield of ethanol, the main fermentation product. The destruction of phytic substances enriched the wort with mineral nutrition and activated the physiological activity of yeast cells. It also led to a slight increase in ethanol yield during the fermentation of the wheat and corn wort (by 2.3–2.4 and 1.5–1.7%, respectively) (Table 4). The ethanol concentration in the mash varied due to a higher starch content in corn (Tables 1 and 4).
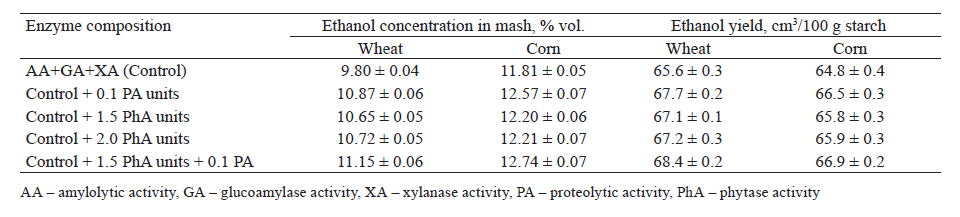
The metabolic processes in yeast cells improved on the media treated with amylolytic, xylanase, and proteolytic enzymes, as well as a complex of enzymes with phytases. This improvement contributed to a complete fermentation of carbohydrates and an increase in ethanol yield, with a simultaneous decrease in associated metabolites (Table 4, Figs. 4 and 5).
The highest concentration of ethanol generated in the wort treated with a full complex of enzymes was 11.15% in the wheat mash and 12.74% in the corn mash. The ethanol yield from the fermentation of wheat and corn wort increased by 3.2 and 2.6%, respectively, when treated with proteases and by 4.3 and 3.2%, respectively, when treated with proteases and phytase (Table 4).
The rise in ethanol synthesis in the experimental samples was probably associated with an improved conversion of polymers of grain wort and its enrichment with assimilable amine nitrogen. Another reason was a release of macro- and microelements that was vital for yeast cells. These findings were consistent with a number of previous studies [2, 7, 25–27].
CONCLUSION
We found almost no changes in the concentration of phosphates during the fermentation of phytases-treated wort, with a slight increase by the end of the process. It was probably caused by the continuing biocatalytic hydrolysis of phytic substances and the release of phosphorus, as well as autolytic processes in the cell. The control samples (without phytolytic enzymes) had a significantly lower residual content of phosphates in the wheat and corn mash (2.4–2.6 and 4.3–5.1 times, respectively).
Our results confirmed that nitrogen and phosphorus nutrition played a regulatory role in the generation and metabolism of ethanol yeast. The catalytic action of phytases and proteases ensured the accumulation of easily assimilable phosphates, minerals, and amino acids in the wort. Also, it intensified the growth of yeast cells and increased the rate of carbohydrate consumption. Finally, it decreased the formation of side metabolites 1.7–1.9 times, mainly due to higher and aromatic alcohols. At the same time, the Saccharomyces cerevisiae 985-T yeast synthesized ethanol, whose yield increased by 1.5–4.3%, depending on the type of grain and enzyme complex. The greatest effect was achieved by a full complex of enzymes (carbohydrase, protease, and phytase).
Contribution
The authors were equally involved in writing the manuscript and are equally responsible for plagiarism.CONFLICTS OF INTEREST
The authors declare that there is no conflict of interest.FUNDING
The study was subsidized by the Program of Fundamental Scientific Research of the State Academies of Sciences.REFERENCES
- Zueva NV, Agafonov GV, Korchagina MV, Dolgov AN. Effect of enzyme preparations on the main parameters of products in the development of processing technology of concentrated wort on ethanol. Proceedings of the Voronezh State University of Engineering Technologies. 2017;79(2):191–197. (In Russ.). https://doi.org/10.20914/2310-1202-2017-2-191-197.
- Serba EM, Abramova IM, Rimareva LV, Overchenko MB, Ignatova NI, Grunin EA. Influence of enzymatic preparations on technological parameters of grain wort and quality of alcohol. Beer and beverages. 2018;(1):50–54. (In Russ.). https://doi.org/10.24411/2072-9650-2018-00002.
- Zueva NV, Agafonov GV, Korchagina MV, Dolgov AN, Chusova AE. Selection of enzyme preparations and temperature-time regimes of water-heat and enzymatic treatment in the development of complex technology of processing of grain raw materials. Proceedings of the Voronezh State University of Engineering Technologies. 2019;81(1):112–119. (In Russ.). https://doi.org/10.20914/2310-1202-2019-1-112-119.
- Caspeta L, Coronel J, Montes de Oca A, Abarca E, González L, Martínez A. Engineering high-gravity fermentations for ethanol production at elevated temperature with Saccharomyces cerevisiae. Biotechnology and Bioengineering. 2019;116(10):2587–2597. https://doi.org/10.1002/bit.27103.
- Fang L, Wang T, Lamsal B. Use of surfactant and enzymes in dry-grind corn ethanol fermentation improves yield of ethanol and distillers corn oil. Industrial Crops and Products. 2018;111:329–335. https://doi.org/10.1016/j.indcrop.2017.10.031.
- Franco JH, Minteer SD, de Andrade AR. Product analysis of operating an ethanol/O2 biofuel cell shows the synergy between enzymes within an enzymatic cascade. Journal of the Electrochemical Society. 2018;165(9):H575–H579. https://doi.org/10.1149/2.0931809jes.
- Guillaume A, Thorigné A, Carré Y, Vinh J, Levavasseur L. Contribution of proteases and cellulases produced by solid-state fermentation to the improvement of corn ethanol production. Bioresources and Bioprocessing. 2019;6(1). https://doi.org/10.1186/s40643-019-0241-0.
- Benesova K, Belakova S, Mikulikova R, Svoboda Z. Survey of the analytical methods for the phytic acid determination. Kvasny Prumysl. 2013;59(5):127–133. https://doi.org/10.18832/kp2013013.
- Mikulski D, Klosowski G. Phytic acid concentration in selected raw materials and analysis of its hydrolysis rate with the use of microbial phytases during the mashing process. Journal of the Institute of Brewing. 2015;121(2):213–218. https://doi.org/10.1002/jib.221.
- Serba EM, Overchenko MB, Ignatova NI, Belokopytova EN, Rimareva LV. Composition of concentrated grain wort, prepared from various types of grain raw materials. Aktualʹnye voprosy industrii napitkov [Current issues of the beverage industry]. 2018;(2):166–170. (In Russ.).
- Zhulʹkov AYu, Vitol IS, Karpilenko GP. Rolʹ zernovoy fitazy pri proizvodstve i sbrazhivanii rzhanogo susla. Chastʹ i. Issledovanie fitaznogo kompleksa rzhi [The role of grain phytase in the production and fermentation of rye wort. Part I. A study of the phytase complex of rye]. Storage and Processing of Farm Products. 2009;(5):50–55. (In Russ.).
- Sapna, Singh B. Phytase production by Aspergillus oryzae in solid-state fermentation and its applicability in dephytinization of wheat ban. Applied Biochemistry and Biotechnology. 2014;173(7):1885–1895. https://doi.org/10.1007/s12010-014-0974-3.
- Hesampour A, Ranaei O, Malboob MA, Harati J, Mohandes N. Comparison of biochemical properties of recombinant phytase expression in the favorable methylotrophic platforms of Pichia pastoris and Hansenula polymorpha. Progress in Biological Sciences. 2014;4(1):97–111. https://doi.org/10.22059/pbs.2014.50309.
- Raman S, Abdullah N, Boo LJ, Azizi S, Mohamad R. Improvement of phytase biosynthesis by new bacterial isolate, Pediococcuspentosaceus C4/1A via continuous cultivation. Journal of Microbiology, Biotechnology and Food Sciences. 2019;8(5):1118–1124. https://doi.org/10.15414/jmbfs.2019.8.5.1118-1124.
- Tarutina MG, Kashirskaya MD, Lazareva MN, Lapteva AR, Dobrynin VYu, Gordeeva TL, et al. Comparative characteristics of phytases from Citrobacter freundii and Yersinia intermedia expressed in Ogataea polymorpha and Pichia pastoris methylotrophic yeasts. Biotechnology in Russia. 2019;35(6):51–56. (In Russ.). https://doi.org/10.21519/0234-2758-2019-35-6-51-56.
- Kulova FM. Effect of the enzymatic preparation fitasa in diets with different mineral content on zootechnical calves’ indexes. Proceedings of Gorsky State Agrarian University. 2016;53(1):71–76. (In Russ.).
- Lenkova TN, Egorova TA, Menʹshenin IA, Sysoeva IG. Otechestvennaya fitaza v kombikormakh dlya kur-nesushek [Domestic phytase in compound feed for laying hens]. Poultry and Poultry Processing. 2016;(1):37–40. (In Russ.).
- Kryukov VS, Glebova IV, Zinoviev SV. Reevaluation of phytase action mechanism in animal nutrition. Biochemistry (Moscow). 2021;86:S152–S165. https://doi.org/10.1134/S000629792114011X.
- Zyba SJ, Wegmüller R, Woodhouse LR, Ceesay K, Prentice AM, Brown KH, et al. Effect of exogenous phytase added to small-quantity lipid-based nutrient supplements (SQ-LNS) on the fractional and total absorption of zinc from a millet-based porridge consumed with SQ-LNS in young Gambian children: a randomized controlled trial. American Journal of Clinical Nutrition. 2019;110(6):1465–1475. https://doi.org/10.1093/ajcn/nqz205.
- Castro-Alba V, Lazarte CE, Bergenståhl B, Granfeldt Y. Phytate, iron, zinc, and calcium content of common Bolivian foods and their estimated mineral bioavailability. Food Science and Nutrition. 2019;7(9):2854–2865. https://doi.org/10.1002/fsn3.1127.
- Bangar P, Glahn RP, Liu Y, Arganosa GC, Whiting S, Warkentin TD. Iron bioavailability in field pea seeds: Correlations with iron, phytate, and carotenoids. Crop Science. 2017;57(2):891–902. https://doi.org/10.2135/cropsci2016.08.0682.
- Kruger J, Oelofse A, Taylor J, Taylor JRN. Potential for improvement in yeast nutrition in raw whole grain sorghum and maize lager brewing and bioethanol production through grain genetic modification and phytase treatment. Journal of the Institute of Brewing. 2012;118(1):70–75. https://doi.org/10.1002/jib.86.
- Rimareva LV, Overchenko MB, Ignatova NI, Krivova AYu, Serba EM. Generation of alcoholic yeast on media prepared from cereals with different phytate content. Food Industry. 2019;(4):83–85. https://doi.org/10.24411/0235-2486-2019-10042.
- Kovaleva TS, Agafonov GV, Yakovlev AN, Yakovleva SF. Effect of protease and phytase on the physiological state of alcoholic yeast in cultivation. Proceedings of the Voronezh State University of Engineering Technologies. 2019;81(4):98–102. (In Russ.). https://doi.org/10.20914/2310-1202-2019-4-98-102.
- Polyakov VA, Serba EM, Overchenko MB, Ignatova NI, Rimareva LV. The effect of a complex phytase-containing enzyme preparation on the process of rye wort fermentation. Foods and Raw Materials. 2019;7(2):221–228. http://doi.org/10.21603/2308-4057-2019-2-221-228.
- Rimareva LV, Krivova AYu, Shelekhova NV, Overchenko MB, Serba EM. Catalytic features of proteas and fitaz when processing politicals of the trititical-without in manufacture of alcohol. Aktualʹnye voprosy industrii napitkov [Current issues of the beverage industry]. 2019;(3):184–190. (In Russ.). https://doi.org/10.21323/978-5-6043128-4-1-2019-3-184-190.
- Lonnerdal B. Phytic acid-trace element (Zn, Cu, Mn) interactions. International Journal of Food Science and Technology. 2002;37(7):749–758. https://doi.org/10.1046/j.1365-2621.2002.00640.x.
- De Carli L, Schnitzler E, Ionashiro M, Szpoganicz B, Rosso ND. Equilibrium, thermoanalytical and spectroscopic studies to characterize phytic acid complexes with Mn(II) and Co(II). Journal of the Brazilian Chemical Society. 2009;20(8):1515–1522. https://doi.org/10.1590/S0103-50532009000800019.
- Lee K-M, Kang H-S, Yun C-H, Kwak H-S. Potential in vitro protective effect of quercetin, catechin, caffeic acid and phytic acid against ethanol-induced oxidative stress in SK-Hep-1 cells. Biomolecules and Therapeutics. 2012;20(5):492–498. https://doi.org/10.4062/biomolther.2012.20.5.492.
- Kumar K, Venkatraman V, Bruheim P. Adaptation of central metabolite pools to variations in growth rate and cultivation conditions in Saccharomyces cerevisiae. Microbial Cell Factories. 2021;20(1). https://doi.org/10.1186/s12934-021-01557-8.
- Meng L, Liu H-L, Lin X, Hu X-P, Teng K-R, Liu S-X. Enhanced multi-stress tolerance and glucose utilization of Saccharomyces cerevisiae by overexpression of the SNF1 gene and varied beta isoform of Snf1 dominates in stresses. Microbial Cell Factories. 2020;19(1). https://doi.org/10.1186/s12934-020-01391-4.
- Zhang Y, Lin Y-H. Metabolic flux analysis of Saccharomyces cerevisiae during redox potential–controlled very high-gravity ethanol fermentation. Biotechnology and Applied Biochemistry. 2020;67(1):140–147. https://doi.org/10.1002/bab.1861.
- De Stefano S, Giuffre O, Milea D, Rigano C, Sammartano S. Speciation of phytate ion in aqueous solution. Non covalent interactions with biogenic polyamines. Chemical Speciation and Bioavailability. 2003;15(2):29–36. https://doi.org/10.3184/095422903782775235.
- Kumar V, Sinha AK, Makkar HPS, Becker K. Dietary roles of phytate and phytase in human nutrition: A review. Food Chemistry. 2010;120(4):945–959. https://doi.org/10.1016/j.foodchem.2009.11.052.
- Zhang YY, Stockmann R, Ng K, Ajlouni S. Revisiting phytate-element interactions: implications for iron, zinc and calcium bioavailability, with emphasis on legumes. Critical Reviews in Food Science and Nutrition. 2020. https://doi.org/10.1080/10408398.2020.1846014.
- Nielsen AVF, Tetens I, Meyer AS. Potential of phytase-mediated iron release from cereal-based foods: A quantitative view. Nutrients. 2013;5(8):3074–3098. https://doi.org/10.3390/nu5083074.
- Yu S, Cowieson A, Gilbert C, Plumstead P, Dalsgaard S. Interactions of phytate and myo-inositol phosphate esters (IP1–5) including IP5 isomers with dietary protein and iron and inhibition of pepsin. Journal of Animal Science. 2012;90(6):1824–1832. https://doi.org/10.2527/jas.2011-3866.
- Tsurikova NV, Nefedova LI, Kostyleva EV, Zvenigorodskii VI, Yasinovskii VG, Voeikova TA, et al. Selection of a potent Bacillus licheniformis strain producing thermostable α-amylase. Applied Biochemistry and Microbiology. 2002;38(5):427–432. https://doi.org/10.1023/A:1019960216770.
- Rimareva LV, Tsurikova NV, Kostyleva EV, Sereda AS. Recombinant mycelial strain aspergillus awamori producer of enzymatic complex of glucoamylase and xylanase. Patent RU 2457246C1. 2012.
- Serba EM, Sokolova EN, Borshcheva YuA, Rimareva LV, Overchenko MB. The effect of chemical mutagenesis on physiological characteristics and productivity of Aspergillus oryzae. Mycology and Phytopathology. 2018;52(1):49–54. (In Russ.).
- Rimareva LV, Overchenko MB, Ignatova NI, Serba EM, Martynenko NN. Features of selected races of alcohol yeast with osmophilic and thermotolerant properties. Storage and Processing of Farm Products. 2016;(10):29–34. (In Russ.).
- Instruktsiya po tekhnokhimicheskomu i mikrobiologicheskomu kontrolyu spirtovogo proizvodstva [Instructions for the technochemical and microbiological control of alcohol production]. Moscow: DeLi print; 2007. 479 p. (In Russ.).
- Serba EM, Overchenko MB, Rimareva LV. Biotekhnologicheskie osnovy mikrobnoy konversii kontsentrirovannogo zernovogo susla v ethanol [Biotechnological principles of microbial conversion of concentrated grain wort into ethanol]. Moscow: Biblio-Globus; 2017. 120 p. (In Russ.). https://doi.org/10.18334/9785950050169.
