

Abstract
Introduction. Traditionally, mammalian adipose tissue is divided into white (white adipose tissue – WAT) and brown (brown adipose tissue – BAT). While the functions of WAT are well known as the triglyceride depot, the role of BAT in mammalian physiology has been under close investigation. The first description of the role of BAT in maintaining thermogenesis dates back to 1961. This article offers a review of structural and functional specificity of white, beige and brown adipose tissue.Results and discussion. The differences and descriptions of adipocytes and their impact on the maintenance of the main functions of the mammalian body are described in this manuscript. In particular, thermogenesis, stress response, obesity, type II diabetes. In addition to WAT and BAT, an intermediate form was also detected in the body – beige fat (BeAT or Brite). The opposite opinions regarding the presence of three types of adipose tissue in the human and animal bodies are presented. Studies on the identification of uncoupling proteins 1 and 3 and their role in the transformation of white fat into beige/brown are considered. Basically, the data on the factors of endogenous and exogenous nature on their formation are given on the example of the human body.
Conclusion. With an abundance of publications on the keywords: “white, brown fat”, these studies, in the overwhelming majority, are devoted to the role of these fats in the formation of human thermogenesis, the assessment of the impact on obesity. Pigs have also been suggested to lack functional BAT, which is a major cause of neonatal death in the swine industry, therefore the focus on investigating role of different types of adipose tissue in pigs seems very promising in order to understand whether there is a compensating mechanism of thermogenesis.
Keywords
Fat, beige and brown adipocytes, uncoupling protein, thermogenesis, adipocyte, animal health, livestockINTRODUCTION
The adipose tissue tissue is a form of connective tissue, the cells of which are filled with a fat drop [1]. Adipose tissue is distributed throughout the body and plays the key role in the body energy homeostasis as a lipid reservoir. Moreover, adipocytes are endocrinologically active, especially visceral [2, 3]. The content of adipose tissue and its lipid composition is strongly depended on biological species, diet, climate, etc.
Historically, adipose tissue of mammals has been divided into two types, white adipose tissue (WAT) and brown adipose tissue (BAT) based on its visible different color, as well as on its different physiological functions [4]. Anatomically, WAT presented in two major depots, subcutaneous and visceral around internal organs and comprises the largest adipose tissue volume in most mammals [2, 4, 5]. WAT is specialized in handling fatty acids and triglycerides (TGs) and critical for energy storage, endocrine communication, and insulin sensitivity [4, 5]. In contrast, amount of BAT is strongly lower. BAT participates in nonshivering thermogenesis and largely present in mammals postnatally and during hibernation [2, 4]. Although BAT is readily observed in both infant and adult mammals, BAT is gradually replaced by WAT with aging [2]. Beige adipose (BeAT) tissue is the third type of adipose tissue and is a result of “browning” of WAT, when brown-like adipocytes appear at anatomical sites characteristic of WAT [6]. It is also called Brite (brown-in-white) [7, 8]. Originally, BeAT was observed to arise in response to cold exposure; however, such factors, as diet, physical activity, pre-and probiotics, pharmaceutical and plantbased substances, etc. can also induce “beigeing” or “browning” of WAT [9].
Health status, survival of offspring, quality of pig processing products are strongly depended on proportion of all three types of adipose tissue. This article offers a review of structural and functional specificity of white, beige and brown adipose tissue, as well as possibility of its formation and divergence in pigs.
RESULTS AND DISCUSSION
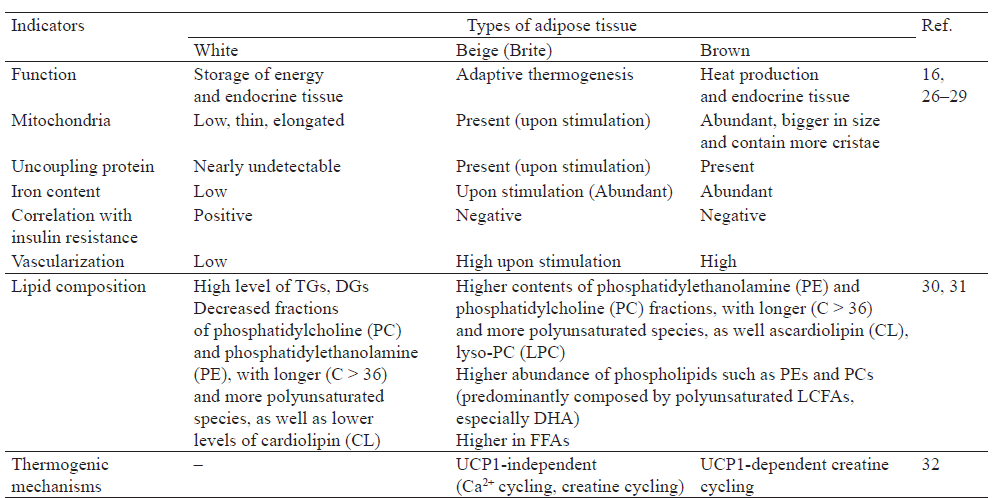
Characteristics of white, beige and brown adipose tissue. The general classification of adipose tissue is based on its color, which is corresponded to lipid content, mitochondrial density and vascularization (Fig. 1). White adipocytes contain a single lipid droplet occupying approximately 90% of the cell space [9]. Brown adipose tissue (BAT) is highly vascularized, brown adipocytes contain a large number of mitochondria, but lipid droplet is smaller and presented in multiple vacuoles [9, 10]. Beige adipocytes display characteristics of both brown and white fat cells, the content of mitochondria is higher than in white adipose tissue (WAT), lipid droplet is not single, but bigger than in BAT [9].
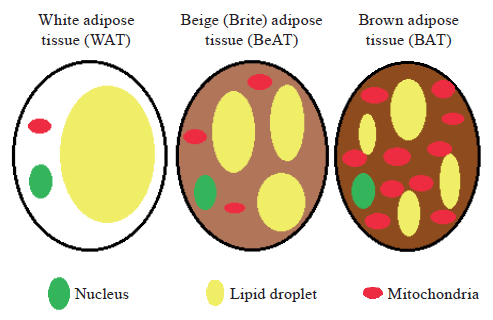
Canonically, adipocytes are different in the origin and thought to arise from the de novo differentiation of precursor cells, particularly, white adipose stem cells originate from Myf5 (myogenic regulatory factor) negative progenitors, whereas brown adipose stem cells originate from myogenic lineage and express Myf5 [12, 13]. Othervise, adipose precursors are heterogeneous, and the exact precursor population for adipogenesis may depend on the sex, location, age, or proadipogenic stimulus [14]. However, more than 95% of the precursors in brown fat are labeled with Myf5-Cre [15].
Beige adipocytes appears in WAT during white to brown transdifferentiation or can arise from adipogenic precursor cells in WAT through de novo differentiation. Additionally, preadipocytes in WAT give rise to mature white adipocytes with the potential to become brite adipocytes at a later point of time, which confirmed that after repeated stimuli most brite adipocytes localize to the same patches within the depot after the first stimuli. Such microenvironment factor, as density of vascularization, the types of stromal-vascular cells in the adipocyte’s vicinity, the composition of the extracellular matrix, and the local innervation could also influence on this process [12, 16, 17].
Different morphology of adipose tissues types corresponds to other divergences (Table 1). WAT mainly maintains energy homeostasis and stores energy in the form of TGs, which are broken down into FFA and glycerol with following catabolism to generate energy when energy demands [10]. It also offers mechanical protection for muscles and internal organs and has an important role in conservation of body temperature [18].
Thus, subcutaneous WAT acts as a shock absorber, providing padding at various anatomic sites, whereas omental WAT is one of the visceral adipose tissue depots, surrounding and protecting inner organs from physical injury [19]. Subcutaneous WAT is more prone to expansion and represents a physiological buffer for excess energy intake during times of limited energy expenditure. When this storage capacity is exceeded, fat accumulates ectopically in areas outside the subcutaneous WAT [19, 20]. Some WAT has only biomechanical function, such as infrapatellar adipose tissue, which preserves even upon extreme starvation [21].
WAT, especially visceral, is also known as an endocrine organ producing adipokines, involved in lipid metabolism or transport, immune system, regulation of pressure, blood coagulation, glycemic homeostasis, angiogenesis, etc. [18, 22]. Adipose tissue also expresses receptors for most of these factors that are implicated in the regulation of many processes, including food intake, energy expenditure, metabolism homeostasis, as well as immunity and blood pressure homeostasis.
Both visceral fat and subcutaneous adipose tissue produce unique profile of adipocytokines, but visceral fat appears to be more active [23]. Excess of WAT is strongly correlated with obesity and insulin resistance [24]. Exceeded fat accumulation in areas outside the subcutaneous WAT, such as lipid accumulation in ectopic tissues (liver, skeletal muscle, and heart) as well as in the visceral depots lead to local inflammation, metabolic disorders and obesity-driven insulin resistance (IR) in WAT, liver, and skeletal muscle [25].
Brown adipose tissue was identified as a thermogenic organ in 1961, in 1978 BAT was shown to be the major site of thermoregulatory non-shivering thermogenesis [33]. However, beneficial effects of BAT could be also explained with its endocrine role through the release of endocrine factors, especially under conditions of thermogenic activation, such insulin-like growth factor I, interleukin-6, or fibroblast growth factor-21, which improve glucose tolerance and insulin sensitivity mainly by influencing hepatic and cardiac function [34]. Comparatively large amounts of BAT are present in the newborns, and then reduced during aging [35].
There is a general proposal that brown adipose tissue is rapidly lost postnatally, the implication being that this process is normally concluded within the first (few) years of life, and that humans later in life do not possess more than vestigical amounts of brown adipose tissue [36].
BAT contains a lot of mitochondria, free fatty acids serve as substrates for lipid oxidation and as potent activators of the mitochondrial uncoupling protein 1 (UCP1), the crucial trans-membrane protein that catalyzes heat production at the mitochondrial level [37]. UCP1 is the only memberable to translocate protons through the inner membrane of brown adipocyte mitochondria, uncouples respiration from ATP synthesis, and therefore provokes energy dissipation in the form of heat while, also stimulating high levels of fatty acid oxidation.
UCP1 homologs were identified but they are biochemically and physiologically different from UCP1 [38]. The biochemical activities and biological roles of the identified UCP2 and UCP3 are poorly understood [39]. UCP2, widely expressed in tissues and cell types, could have particular importance in the regulation in lipid metabolism and contribute to resting metabolic rate, fat oxidation, while UCP3 could transport protons with a rate comparable to UCP1 [40–42]. Thus, it was shown that UCP3 is highly abundant in BAT and the sensitivity of the protein expression to temperature is similar to that of UCP1 [43]. It was also revealed that UCP3, expressed in both skeletal muscle and brown adipose tissue, may act as an inducible thermogenin in vivo and could indirectly mediate thermogenesis by increasing fatty acid oxidation and metabolite transport [44]. However, UCP2 and UCP3 are not generally responsible for adaptive thermogenesis, but nonetheless they may be significantly thermogenic when fully activated by endogenous or exogenous effectors [45].
BAT has negative correlation with obesity and insulin resistance, increasing BAT mass could improve glucose metabolism and metabolic health [46]. Thus, it was estimated that 50 g of BAT can burn as much as 20% of daily energy intake; therefore even though the BAT depots are present in small amounts, the activated tissue has the potential to substantially contribute to energy expenditure. In addition to using lipids, BAT also displays a very high rate of glucose uptake under cold exposure, glucose uptake increases by 12-folds, dissipating energy as a function of increased blood flow [47].
Beige – brown in white or brite (BeAT) – an intermediate type of fat, which is similar functionally to brown fat – it has a high thermogenicity and contains a significant number of mitochondria. Nevertheless, beige adipocytes may secrete certain factors that affect WAT function, systemic metabolism or both, has negative correlation with obesity and insulin resistance and appears upon the stimulation into WAT [29, 48, 49]. BeAT plays the key role in adaptive thermogenesis, subcutaneous WAT is particularly prone to browning [50]. Thermogenic capacity of beige fat cells depends on the presence of UCP1 [51].
Localization of beige and brown adipose tissue. Distribution of BAT and BeAT is different, localization is various in species; it is most studied in humans and rodents (Fig. 2). In human infants, BAT is located in interscapular and peri-renal areas, while in adults smaller BAT depots are located in the anterior cervical, supraclavicular, axillary, peri-aortic, paravertebral and suprarenal regions, while beige fat signature could be formed in supraclavicular, abdominal visceral and subcutaneous fat depots [32, 48, 52–57]. However, distribution of human BAT could be wider. Visceral BAT includes perivascular (aorta, common carotid artery, brachiocephalic artery, paracardiac mediastinal fat, epicardial coronary artery and cardiac veins, as well as internal mammary and intercostal artery and vein), periviscus (heart, trachea and major bronchi at lung hilum, esophagus, greater omentum, transverse mesocolon) and around solid organs (thoracic paravertebral, pancreas, kidney, adrenal, liver, hilum of spleen). Subcutaneous BAT includes depots lying between the anterior neck muscles and in the supraclavicular fossa, under the clavicles, in the axilla, in the anterior abdominal wall, and in the inguinal area [58]. Beige fat is could be also detected in cervical, parasternal, supraclavicular, para- and prevetebral areas [59].
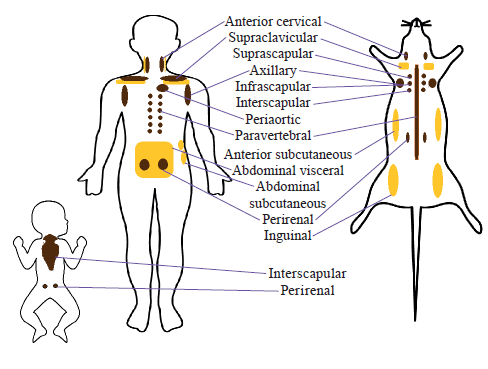
In mouse BAT is located in anterior cervical, supraclavicular, axillary, interscapular, infrascapular, paravertebral and perirenal areas, while BeAT – in anterior subcutaneous WAT, supraclavicular WAT and inguinal WAT [32, 54, 56]. The main differences between human and mouse adipocytes are defined. Human BAT are dispersed and represented a mix of white, classical brown, and recruitable brite adipocytes, while murine the main BAT depots are in well-defined anatomic sites and homogeneously composed of brown adipocytes [60].
The ways of “browning” of white adipose tissue. Beige adipocytes were originally observed to arise in response to cold; however, studies have since identified that diet, exercise, pre- and probiotics, pharmaceutical agents, numerous plant-based bioactives, and even adipokines, can also induce “beigeing” or “browning” of WAT [61]. Thus, “beigeing” or “browning” of WAT could be caused by β-3 adrenergic receptor agonists, (CL 316243, BRL 26830A), short-chain fatty acids (acetate), dietary factors and organic compounds (capsaicin (and related capsinoids), plant-produced resveratrol, plant-derived berberine (BBR), fish oil, decaffeinated green tea extract, cinnamon extract, ginsenoside Rb1, curcumin, quercetin, ginger extract), nuclear receptors and ligands (farnesoid X receptor, liver X receptors), microRNAs (miRNA-32, miRNA-455), drug agents (Thiazolidinediones, Prostaglandin E2, Gleevec, Beta-lapachone, Slit 2 derived secretory product, Artepillin C, Adrenomedullin 2), inflammatory factors (IL-6, IL-4, IEX-1), hormonal factors (thyroid hormones, Glucagon-like peptide 1, leptin, melatonin, natriuretic peptides), genetic factors (PTEN, Cox2, Foxc2, folliculin, Gq, TGF-β/Smad3), batokines (FGF21, apelin), exercise, PPAR agonists (rosiglitazone,WY14643), bone morphogenetic proteins (BMP7, BMP4), metabolites (lactate, β-hydroxybutyrate, beta-aminoisobutyric acid (BAIBA), retinoic acid), bariatric surgery (physical reconstruction of the gastrointestinal tract) [10, 49].
Nevertheless, the most studied factor is cold exposure. Thus, cold is sensed by the skin and central signals result in increased noradrenaline release via sympathetic neurons and subsequent stimulation of various subtypes of β-adrenergic receptors (ADRBs, mainly subtype β3) and downstream cyclic adenosine monophosphate signaling, leading to the proliferation of brown adipocytes and activation of lipolysis and/or of thermogenesis [62–64]. However, cold-induced BAT from adult human neck area consists of classical brown adipocytes, as well as activated thermogenic fat in the supraclavicular region is composed of both classical brown and beige adipocytes [65].
Nutritional induction is also studied. WAT content is influenced by n-3 PUFA, polyphenols, vitamin D, vitamin E, vitamin A, carotenoids, BAT – by PUFA, especially n-3 PUFA, bile acids, BeAT – by amino acid restriction, capsaicin, bile acids, n-3 PUFA, retinoic acid [27]. Low protein diet results in activation of brown adipose tissue, as well as sucrose intake increase BAT activity. Some of the diet-derived small molecules increase BAT activity and browning of WAT, such as acetic, butyric and succinic acids, ketone bodies. Consumption of chilli peppers (capsaicin, non-pungent capsinoids), olive oil (oleic acid), green tea (catechins), raspberry (RB-ketone), grapes (resveratrol), fish (PUFAs) also stimulate BAT activation and browning [66].
Cannon and Nedergaard described the mechanism of transformation controlled by hypothalamus [67]. In the experiment on obese ob/ob mice consumed cafeteria diet (overfed) an activation of brown fat was observed – diet-induced thermogenesis (DIT) [33]. In general, macronutrient content of meals (carbohydrate, fat, protein amount and type) and dietary bioactive compounds (capsaicin and capsinoids; tea, caffeine and catechins; menthol; conjugated linoleic acid, casein protein, curcumin, garlic powder, procyanidinrich extracts from black soybean seed, resveratrol and extracts from ginger family plants, etc.) could affect BAT and browning process [68]. Interestingly, that gut microbiota could contribute to upregulation of thermogenesis in the cold environment [69]. Zhang et al. reported that Caulis spatholobi can activate brown adipose tissue and modulate the composition of gut microbiota, which is linked with normalization of thermogenesis during cooling [70].
Most of the research describe functions and role of WAT, BAT and BeAT in human body. Not many scientists deal with adipogenic features of white, beige, and brown adipose tissues in other mammalian, domestic farm animals in particular.
Adipose tissues types in pigs. Database search sciencedirect.com showed that according to the keywords “brown fat, white fat”, the system issues 3475 publications for 2020, the number of publications has doubled in 10 years. 6557 scientific papers were published in 2020 for the keywords “brown fat, browning”. When the keyword “pig” is added to these keywords, the number of publications is reduced to 356.
The analysis of these publications showed that the main scientific interest is directed on modification of fatty acid composition, but not on the study of the fat types and their distribution, although the directed modification of fatty acid profile is of considerable interest, taking into account correlation of the fatty acids amount with long chain activity of mitochondrial uncoupling protein 1 (UCP1), the activation of the mitochondria of brown fat and non-shivering thermogenesis.
Uncoupling protein 1 (UCP1), is a unique mitochondrial membranous protein devoted to adaptive thermogenesis, a specialized function performed by brown adipocytes [38]. The restricted interest to BAT and BeAT in pigs is explained that pigs (Suidae) have a predominantly tropical distribution and lost functional UCP1 in a genetic event that eliminated exons 3–5 ~ 20 million years ago [71–73]. They consequently have also been suggested to lack functional BAT, which is a major cause of neonatal death in the swine industry [71].
Despite these inconsistent findings, some pig breeds, such as the Tibetan pig found on the Qinghai-Tibetan plateau and the Min pig living in Northeast China, are well recognized to be cold resistant, and WAT browning was induced after cold exposure as well as UCP3 expression was significantly increased. Coldresistant pig breeds (eight dominant pig breeds found across China) have evolved a novel mechanism involving UCP3 in beige adipocytes as the primary thermogenic mechanism, challenging the orthodoxy based on studies of mice that only UCP1 may act as a significant source of thermogenic heat [71]. Pigs do not have BAT, but beige adipocytes were found in inguinal subcutaneous WAT, axillary sWAT and perirenal fat from acute coldstimulated cold-tolerant pig breeds in China, including Tibetan pigs and Min pigs (Fig. 3) [74]. Differentiated beige cells were also observed in subcutaneous fat of Tibetan pigs [71].
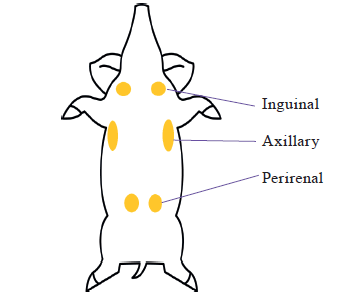
As beige adipocytes were observed at least in cold stimulated adipose tissues from cold-resistant pigs, UCP1-independent non-shivering thermoregulation might be justified with temperature maintenance in pigs or UCP3-dependent thermogenesis in beige adipocytes as a key evolutionary response in cold-adapted pig lineages [71, 74]. The studies in this area are important, especially concerning neonatal death in the swine industry and expanding the geography of pig farming.
CONCLUSION
White, brown, and beige/brite adipose tissues are considered mainly from the point of view of human health, paying special attention to their role in obesity and type II diabetes. Mechanisms and tools of white adipose tissue browning are intensively studied, as well as brown and beige/brite adipose tissues localization and features in different species. The phenotypic and genotypic study of various breeds of pigs in different conditions of housing, taking into account climatic zones, will help reveal the main qualitative characteristics of fat.
The new knowledge about beige adipose tissue with some similarity to brown, which is characteristic of the neonatal period and almost disappears in the adult body and has a thermogenic function, opens up new opportunities for the formation of qualitatively new characteristics of pig adipose tissue.
Using the knowledge about the influence of a number of endo- and exogenous factors on the formation of adipose tissue (white, beige, brown), it will be possible to control the molecular mechanisms of adipocyte differentiation to obtain not only high-quality pork fat, but also meat products, as well as to expand the geography of pig breeding.
Contribution
The authors were equally involved in writing the manuscript and are equally responsible for plagiarism.CONFLICTS OF INTEREST
The authors state that there is no conflict of interest.FUNDING
This work was financially supported by the Russian Science Foundation (RSF) (project No. 21-76-20032).REFERENCES
- Rosen ED, Spiegelman BM. What we talk about when we talk about fat. Cell. 2014;156(1–2):20–44. https://doi.org/10.1016/j.cell.2013.12.012.
- Choe SS, Huh JY, Hwang IJ, Kim JI, Kim JB. Adipose tissue remodeling: Its role in energy metabolism and metabolic disorders. Frontiers in Endocrinology. 2016;7. https://doi.org/10.3389/fendo.2016.00030.
- McNamara JP, Huber K. Metabolic and endocrine role of adipose tissue during lactation. Annual Review of Animal Biosciences. 2018;6:177–195. https://doi.org/10.1146/annurev-animal-030117-014720.
- Richard AJ, White U, Elks CM, Stephens JM, Feingold KR, Anawalt B, et al. Adipose tissue: Physiology to metabolic dysfunction. In: Feingold KR, Anawalt B, Boyce A, Chrousos G, de Herder WW, Dhatariya K, et al., editors. Endotext. South Dartmouth: MDText.com; 2000.
- Scheja L, Heeren J. Metabolic interplay between white, beige, brown adipocytes and the liver. Journal of Hepatology. 2016;64(5):1176–1186. https://doi.org/10.1016/j.jhep.2016.01.025.
- Giralt M, Villarroya F. White, brown, beige/brite: Different adipose cells for different functions? Endocrinology. 2013;154(9):2992–3000. https://doi.org/10.1210/en.2013-1403.
- Wang H, Willershäuser M, Karlas A, Gorpas D, Reber J, Ntziachristos V, et al. A dual Ucp1 reporter mouse model for imaging and quantitation of brown and brite fat recruitment. Molecular Metabolism. 2019;20:14–27. https://doi.org/10.1016/j.molmet.2018.11.009.
- Chu D-T, Gawronska-Kozak B. Brown and brite adipocytes: Same function, but different origin and response. Biochimie. 2017;138:102–105. https://doi.org/10.1016/j.biochi.2017.04.017.
- Pilkington A-C, Paz HA, Wankhade UD. Beige adipose tissue identification and marker specificity-overview. Frontiers in Endocrinology. 2021;12. https://doi.org/10.3389/fendo.2021.599134.
- Mulya A, Kirwan JP. Brown and beige adipose tissue: Therapy for obesity and its comorbidities? Endocrinology and Metabolism Clinics of North America. 2016;45(3):605–621. https://doi.org/10.1016/j.ecl.2016.04.010.
- Cheng L, Wang J, Dai H, Duan Y, An Y, Shi L, et al. Brown and beige adipose tissue: a novel therapeutic strategy for obesity and type 2 diabetes mellitus. Adipocyte. 2021;10(1):48–65. https://doi.org/10.1080/21623945.2020.1870060.
- Maurer S, Harms M, Boucher J. The colorful versatility of adipocytes: white-to-brown transdifferentiation and its therapeutic potential in humans. FEBS Journal. 2021;288(12):3628–3646. https://doi.org/10.1111/febs.15470.
- Panina YA, Yakimov AS, Komleva YK, Morgun AV, Lopatina OL, Malinovskaya NA, et al. Plasticity of adipose tissue-derived stem cells and regulation of angiogenesis. Frontiers in Physiology. 2018;9. https://doi.org/10.3389/fphys.2018.01656.
- Hepler C, Vishvanath L, Gupta RK. Sorting out adipocyte precursors and their role in physiology and disease. Genes and Development. 2017;31(2):127–140. https://doi.org/10.1101/gad.293704.116.
- Sanchez-Gurmaches J, Guertin DA. Adipocyte lineages: Tracing back the origins of fat. Biochimica et Biophysica Acta (BBA) – Molecular Basis of Disease. 2014;1842(3):340–351. https://doi.org/10.1016/j.bbadis.2013.05.027.
- Wang WS, Seale P. Control of brown and beige fat development. Nature Reviews Molecular Cell Biology. 2016;17(11):691–702. https://doi.org/10.1038/nrm.2016.96.
- Rosenwald M, Wolfrum C. The origin and definition of brite versus white and classical brown adipocytes. Adipocyte. 2014;3(1):4–9. https://doi.org/10.4161/adip.26232.
- Fonseca-Alaniz MH, Takada J, Alonso-Vale MI, Lima FB. Adipose tissue as an endocrine organ: from theory to practice. Jornal de Pediatria. 2007;83(5):S192–S203. https://doi.org/10.2223/JPED.1709.
- Alkhouli N, Mansfield J, Green E, Bell J, Knight B, Liversedge N, et al. The mechanical properties of human adipose tissues and their relationships to the structure and composition of the extracellular matrix. American Journal of Physiology. Endocrinology and Metabolism. 2013;305(12):E1427–E1435. https://doi.org/10.1152/ajpendo.00111.2013.
- Chait A, den Hartigh LJ. Adipose tissue distribution, inflammation and its metabolic consequences, including diabetes and cardiovascular disease. Frontiers in Cardiovascular Medicine. 2020;7. https://doi.org/10.3389/fcvm.2020.00022.
- Zwick RK, Guerrero-Juarez CF, Horsley V, Plikus MV. Anatomical, physiological, and functional diversity of adipose tissue. Cell Metabolism. 2018;27(1):68–83. https://doi.org/10.1016/j.cmet.2017.12.002.
- Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. Journal of Clinical Endocrinology and Metabolism. 2004;89(6):2548–2556. https://doi.org/10.1210/jc.2004-0395.
- Coelho M, Oliveira T, Fernandes R. Biochemistry of adipose tissue: an endocrine organ. Archives of Medical Science. 2013;9(2):191–200. https://doi.org/10.5114/aoms.2013.33181.
- Gómez-Hernández A, Beneit N, Díaz-Castroverde S, Escribano Ó. Differential role of adipose tissues in obesity and related metabolic and vascular complications. International Journal of Endocrinology. 2016;2016. https://doi.org/10.1155/2016/1216783.
- Longo M, Zatterale F, Naderi J, Parrillo L, Formisano P, Raciti GA, et al. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. International Journal of Molecular Sciences. 2019;20(9). https://doi.org/10.3390/ijms20092358.
- Park A, Kim WK, Bae K-H. Distinction of white, beige and brown adipocytes derived from mesenchymal stem cells. World Journal of Stem Cells. 2014;6(1):33–42. https://doi.org/10.4252/wjsc.v6.i1.33.
- Bargut TCL, Souza-Mello V, Aguila MB, Mandarim-de-Lacerda CA. Browning of white adipose tissue: lessons from experimental models. Hormone Molecular Biology and Clinical Investigation. 2017;31(1). https://doi.org/10.1515/hmbci-2016-0051.
- Cedikova M, Kripnerová M, Dvorakova J, Pitule P, Grundmanova M, Babuska V, et al. Mitochondria in white, brown, and beige adipocytes. Stem Cells International. 2016;2016. https://doi.org/10.1155/2016/6067349.
- Villarroya F, Cereijo R, Gavaldà-Navarro A, Villarroya J, Giralt M. Inflammation of brown/beige adipose tissues in obesity and metabolic disease. Journal of Internal Medicine. 2018;284(5):492–504. https://doi.org/10.1111/joim.12803.
- Leiria LO, Tseng Y-H. Lipidomics of brown and white adipose tissue: Implications for energy metabolism. Biochimica et Biophysica Acta (BBA) – Molecular and Cell Biology of Lipids. 2020;1865(10). https://doi.org/10.1016/j.bbalip.2020.158788.
- Schweizer S, Liebisch G, Oeckl J, Hoering M, Seeliger C, Schiebel C, et al. The lipidome of primary murine white, brite, and brown adipocytes – Impact of beta-adrenergic stimulation. PLoS Biology. 2019;17(8). https://doi.org/10.1371/journal.pbio.3000412.
- Ikeda K, Maretich P, Kajimura S. The common and distinct features of brown and beige adipocytes. Trends in Endocrinology and Metabolism 2018;29(3):191–200. https://doi.org/10.1016/j.tem.2018.01.001.
- Trayhurn P. Origins and early development of the concept that brown adipose tissue thermogenesis is linked to energy balance and obesity. Biochimie. 2017;134:62–70. https://doi.org/10.1016/j.biochi.2016.09.007.
- Villarroya J, Cereijo R, Villarroya F. An endocrine role for brown adipose tissue? American journal of physiology – Endocrinology and metabolism. 2013;305(5):E567–E572. https://doi.org/10.1152/ajpendo.00250.2013.
- Symonds ME. Brown adipose tissue growth and development. Scientifica. 2013;2013. https://doi.org/10.1155/2013/305763.
- Nedergaard J, Bengtsson T, Cannon B. Unexpected evidence for active brown adipose tissue in adult humans. American Journal of Physiology – Endocrinology and Metabolism. 2007;293(2):E444–E452. https://doi.org/10.1152/ajpendo.00691.2006.
- Oelkrug R, Polymeropoulos ET, Jastroch M. Brown adipose tissue: physiological function and evolutionary significance. Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology. 2015;185(6):587–606. https://doi.org/10.1007/s00360-015-0907-7.
- Ricquier D. Uncoupling protein 1 of brown adipocytes, the only uncoupler: a historical perspective. Frontiers in Endocrinology. 2011;2. https://doi.org/10.3389/fendo.2011.00085.
- Ricquier D, Bouillaud F. The uncoupling protein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP. Biochemical Journal. 2000;345(2):161–179. https://doi.org/10.1042/0264-6021:3450161.
- Ricquier, D. Uncoupling protein-2 (UCP2): Molecular and genetic studies. International Journal of Obesity. 1999;23:S38–S42. https://doi.org/10.1038/sj.ijo.0800942.
- Saleh MC, Wheeler MB, Chan CB. Uncoupling protein-2: evidence for its function as a metabolic regulator. Diabetologia. 2002;45(2):174–187. https://doi.org/10.1007/s00125-001-0737-x.
- Pohl EE, Rupprecht A, Macher G, Hilse KE. Important trends in UCP3 investigation. Frontiers in Physiology. 2019;10. https://doi.org/10.3389/fphys.2019.00470.
- Hilse KE, Kalinovich AV, Rupprecht A, Smorodchenko A, Zeitz U, Staniek K, et al. The expression of UCP3 directly correlates to UCP1 abundance in brown adipose tissue. Biochimica et Biophysica Acta (BBA) – Bioenergetics. 2016;1857(1):72–78. https://doi.org/10.1016/j.bbabio.2015.10.011.
- Riley CL, Dao C, Kenaston MA, Muto L, Kohno S, Nowinski SM, et al. The complementary and divergent roles of uncoupling proteins 1 and 3 in thermoregulation. Journal of Physiology. 2016;594(24):7455–7464. https://doi.org/10.1113/JP272971.
- Brand MD, Esteves TC. Physiological functions of the mitochondrial uncoupling proteins UCP2 and UCP3. Cell Metabolism. 2005;2(2):85–93. https://doi.org/10.1016/j.cmet.2005.06.002.
- White JD, Dewal RS, Stanford KI. The beneficial effects of brown adipose tissue transplantation. Molecular Aspects of Medicine. 2019;68:74–81. https://doi.org/10.1016/j.mam.2019.06.004.
- Poher AL, Altirriba J, Veyrat-Durebex C, Rohner-Jeanrenaud F. Brown adipose tissue activity as a target for the treatment of obesity/insulin resistance. Frontiers in Physiology. 2015;6. https://doi.org/10.3389/fphys.2015.00004.
- Harms M, Seale P. Brown and beige fat: development, function and therapeutic potential. Nature Medicine. 2013;19(10):1252–1263. https://doi.org/10.1038/nm.3361.
- Kaisanlahti A, Glumoff T. Browning of white fat: agents and implications for beige adipose tissue to type 2 diabetes. Journal of Physiology and Biochemistry. 2019;75(1). https://doi.org/10.1007/s13105-018-0658-5.
- Keipert S, Jastroch M. Brite/beige fat and UCP1 – Is it thermogenesis? Biochimica et Biophysica Acta (BBA) – Bioenergetics. 2014;1837(7):1075–1082. https://doi.org/10.1016/j.bbabio.2014.02.008.
- Bartesaghi S, Hallen S, Huang L, Svensson P-A, Momo RA, Wallin S, et al. Thermogenic activity of UCP1 in human white fat-derived beige adipocytes. Molecular Endocrinology. 2015;29(1):130–139. https://doi.org/10.1210/me.2014-1295.
- Jung SM, Sanchez-Gurmaches J, Guertin DA. Brown adipose tissue development and metabolism. Handbook of Experimental Pharmacology. 2019;251:3–36. https://doi.org/10.1007/164_2018_168.
- Zoico E, Rubele S, De Caro A, Nori N, Mazzali G, Fantin F, et al. Brown and beige adipose tissue and aging. Frontiers in Endocrinology. 2019;10. https://doi.org/10.3389/fendo.2019.00368.
- Sidossis L, Kajimura S. Brown and beige fat in humans: thermogenic adipocytes that control energy and glucose homeostasis. Journal of Clinical Investigation. 2015;125(2):478–486. https://doi.org/10.1172/JCI78362.
- Peirce V, Carobbio S, Vidal-Puig A. The different shades of fat. Nature. 2014;510(7503):76–83. https://doi.org/10.1038/nature13477.
- Cohen P, Kajimura S. The cellular and functional complexity of thermogenic fat. Nature Reviews Molecular Cell Biology. 2021;22(6):393–409. https://doi.org/10.1038/s41580-021-00350-0.
- Chechi K, Lichtenbelt WM, Richard D. Brown and beige adipose tissues: phenotype and metabolic potential in mice and men. Journal of Applied Physiology. 2018;124(2):482–496. https://doi.org/10.1152/japplphysiol.00021.2017.
- Sacks H, Symonds ME. Anatomical locations of human brown adipose tissue: functional relevance and implications in obesity and type 2 diabetes. Diabetes. 2013;62(6):1783–1790. https://doi.org/10.2337/db12-1430.
- Fenzl A, Kiefer FW. Brown adipose tissue and thermogenesis. Hormone Molecular Biology and Clinical Investigation. 2014;19(1):25–37. https://doi.org/10.1515/hmbci-2014-0022.
- Liu X, Cervantes C, Liu F. Common and distinct regulation of human and mouse brown and beige adipose tissues: a promising therapeutic target for obesity. Protein and Cell. 2017;8(6):446–454. https://doi.org/10.1007/s13238-017-0378-6.
- Pilkington AC, Paz HA, Wankhade UD. Beige adipose tissue identification and marker specificity – overview. Frontiers in Endocrinology. 2021;12. https://doi.org/10.3389/fendo.2021.599134.
- Kuryłowicz A, Puzianowska-Kuźnicka M. Induction of adipose tissue browning as a strategy to combat obesity. International Journal of Molecular Sciences. 2020;21(17). https://doi.org/10.3390/ijms21176241.
- Roth CL, Molica F, Kwak BR. Browning of white adipose tissue as a therapeutic tool in the fight against atherosclerosis. Metabolites. 2021;11(5). https://doi.org/10.3390/metabo11050319.
- Herz CT, Kiefer FW. Adipose tissue browning in mice and humans. Journal of Endocrinology. 2019;241(3):R97–R109. https://doi.org/10.1530/JOE-18-0598.
- Wang S, Yang X. Inter-organ regulation of adipose tissue browning. Cellular and Molecular Life Sciences. 2017;74(10):1765–1776. https://doi.org/10.1007/s00018-016-2420-x.
- Srivastava S, Veech RL. Brown and brite: The fat soldiers in the anti-obesity fight. Frontiers in Zhysiology. 2019;10. https://doi.org/10.3389/fphys.2019.00038.
- Cannon B, Nedergaard J. Brown adipose tissue: Function and physiological significance. Physiological Reviews. 2004;84(1):277–359. https://doi.org/10.1152/physrev.00015.2003.
- Suchacki KJ, Stimson RH. Nutritional regulation of human brown adipose tissue. Nutrients. 2021;13(6). https://doi.org/10.3390/nu13061748.
- Li B, Li L, Li M, Lam SM, Wang G, Wu Y, et al. Microbiota depletion impairs thermogenesis of brown adipose tissue and browning of white adipose tissue. Cell Reports. 2019;26(10):2720–2737. https://doi.org/10.1016/j.celrep.2019.02.015.
- Zhang C, Liu J, He X, Sheng Y, Yang C, Li H, et al. Caulis spatholobi ameliorates obesity through activating brown adipose tissue and modulating the composition of gut microbiota. International Journal of Molecular Sciences. 2019;20(20). https://doi.org/10.3390/ijms20205150.
- Lin J, Cao C, Tao C, Ye R, Dong M, Zheng Q, et al. Cold adaptation in pigs depends on UCP3 in beige adipocytes. Journal of Molecular Cell Biology. 2017;9(5):364–375. https://doi.org/10.1093/jmcb/mjx018.
- Fyda TJ, Spencer C, Jastroch M, Gaudry MJ. Disruption of thermogenic UCP1 predated the divergence of pigs and peccaries. Journal of Experimental Biology. 2020;223(15). https://doi.org/10.1242/jeb.223974.
- Jastroch M, Andersson L. When pigs fly, UCP1 makes heat. Molecular Metabolism. 2015;4(5):359–362. https://doi.org/10.1016/j.molmet.2015.02.005.
- Zhao J, Tao C, Chen C, Wang Y, Liu T. Formation of thermogenic adipocytes: What we have learned from pigs. Fundamental Research. 2021;1(4):495–502. https://doi.org/10.1016/j.fmre.2021.05.004.
