

Аннотация
The increasing volume of consumption of probiotics and functional food products requires determination of standardized criteria for cultures and their exometabolites used in functional products manufacturing. The study was aimed at developing criteria for the estimation and standardization of exometabolites and the colony-forming ability of probiotic strains for functional food production. The work included such microbiological and physicochemical methods as GC-MS, GPC, UV, and FT-IR-spectroscopy. Based on the results of the study, the comparative analysis of the microbiological properties of probiotic Bifidobacterium strains was provided, the fatty acid composition of the cell wall was described, and the physical and chemical study of the exopolymers produced by them was carried out. According to the data of FT-IR- spectroscopy, the characteristic features of the components of the cell wall of Bifidobacterium strains were established. Bifidobacteria form the unique composition of organophosphorus structures of lipoteichoic acids, which determines the adhesive ability of strains. The authors studied the molecular weight distribution of the samples of exometabolites isolated from the nutrient medium after the cultivation of bifidobacteria, under conditions of gel-permeation chromatography. The spectral (UV, FT-IR) characteristics of the produced metabolites and their chromatographic fractions were compared. The fatty acids of the Bifidobacterium cell membrane were analyzed using the GC/MS method. The fatty acids were extracted from bacterial cells with different hydrophobicity with a mixture of chloroform and hexane. It has been established that the hydrophobicity is determined by different contents of unsaturated and branched fatty acids in the bacterial membrane. Hydrophobic bifidobacteria are the only that contain the isopentadecane (isoC15:0) and methyl-tetradecanoic (13Me-C14:0) acids. With the mean hydrophobicity, a high content of the isopalmitic (isoC16:0) and stearic (C18:0) acids was established. Low-hydrophobic strains are characterized by a low content of monounsaturated fatty acids.Ключевые слова
Bifidobacteria, hydrophobicity, exometabolites, lipoteichoic acidsВВЕДЕНИЕ
Probiotics are the promising components of functional food products, as well as of biologically active supplements [1]. The demand for probiotic functional products is growing rapidly due to an increase in the consumer awareness of the effect of food on health and life expectancy.
Most of probiotics are used as the functional food products that are a significant share of the food and pharmaceutical market in the developed countries and displace a lot of traditional medicines from sales. It should be noted that probiotics maintain the normal intestinal microflora, protect the gastrointestinal tract from colonization by pathogenic microorganisms, showing the antagonistic properties to the latter, improve the functioning of the body's immune system, reduce serum cholesterol, and normalize blood pressure. All these contribute to health maintenance [2–4]. It is also proved that some probiotic strains have an anticarcinogenic activity and contribute to the improvement of metabolism and an increase in the nutritional value of food products. From year to year, the list of pathological conditions extends, which requires the use of probiotics both as part of medicines and in the form of functional food products. Probiotics are used to prevent diabetic diarrhea, urogenital diseases, osteoporosis, food allergies, and atopic conditions, as well as to relieve constipation and to reduce hypercholesterolemia [5].
The creation of functional food products while maintaining the adequate number of probiotic microorganisms is a problem, since there are many factors, both during production and when storing the finished product, which affect the viability of probiotic bacteria [6]. However, it is important to estimate the strains themselves selected for specific functional probiotic properties. First of all, this is the possibility of providing colonization resistance, which is caused by the adhesive activity of microorganisms. Probiotic bacteria should have the ability to competitively adhere to the epithelium through glycoconjugate receptors, closing the attachment points for pathogenic microorganisms. Using the example of Caco-2 colonocyte culture, it was shown that living probiotic strains, adhering to the epithelium due to the induction of the synthesis of TM-5 tropomyosin, actin and occlusin, strengthen the cytoskeleton of enterocyte cells. In parallel with this, the MUC-3 gene is stimulated, the phosphorylation of the protein of the intercellular compounds increases, the synthesis is stimulated, the epithelial growth factor receptor is activated, and the synthesis of the polyamines, which play the role of hormone-like substances, increases. As a result, the permeability of intercellular compounds decreases, the mucin synthesis increases, and the epithelial regeneration processes intensifies [7, 8]. These mechanisms, as a whole, increase the resistance of the intestinal epithelium and its protective functions. However, these effects were obtained and studied in vitro using a limited number of probiotic bacteria, whereas in the human body they can be leveled or realized in a low volume therefore approaches are needed to estimate the factors of microorganism adhesion, rather than the process of bacterial adhesion [9]. It was proved that probiotics take part in the formation of free amino acids, organic acids, oligosaccharides, short chain fatty acids, bioactive peptides, and bacteriocins, as well as affect the synthesis of vitamins such as biotin, vitamin K, and the vitamin B complex [10, 11]. Therefore, the estimation of the exometabolites produced by probiotic strains is also urgent and is sought after the producer.
To provide a single scientifically grounded approach to the estimation of the quality and safety of the strains of probiotic microorganisms intended for the production of direct probiotic starters, bacterial concentrates, and the microbial biomass, which is a component of food and biologically active supplements or used to enrich traditional food products, some methodical guidelines were developed [12]. The estimation included a complex of microbiological, biochemical, molecular genetic, and hygienic studies to confirm their safety (harmlessness), as well as the presence of their properties that determine the probiotic effect in the body and are therefore suitable for creating probiotic food products and biologically active food supplements. In this case, it is necessary to take into account a large number of factors and properties of potential strains of microorganisms in order to achieve a further health-saving effect [13]. Classical microbiological methods are routine, time-consuming, cost-intensive and often limitedly characterize the properties of the strain under study. Therefore, when searching for original probiotic cultures meeting the requirements of medical science, dietology, the food industry, and biotechnology, it is promising to use complex approaches to the estimation of their properties. In this regard, the criteria for estimating the functional activity of probiotic strains and their safety, regulated by this methodical document [12], are of special significance. However, this regulatory document does not provide any specific methods for studying the antioxidant activity of bacteria and for determining the composition of exopolysaccharides. The colony-forming abikity of probiotic cultures was proposed to be determined in vitro and in vivo using biological models (erythrocytes in rabbits), whereas it is more informative to study precisely the factors of specific and nonspecific adhesion in a specific strain.
The study was aimed at developing criteria for the estimation and standardization of exometabolites and the colonization potential of the probiotic strains intended for manufacturing functional food products.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
The object of the study was the probiotic strains of Bifidobacterium bifidum 1, B. longum B 379M, and B. breve isolated from commercial preparations. The bifidobacteria were isolated from preparations using a routine bacteriological method. Anaerobic jars (BBL, USA) and gas-generating packets (NPO Novoe Delo, St. Petersburg) were used to create anaerobic conditions. The phenotypic biochemical profile of strains was studied using commercial ANAERO- TEST 23 test systems (Lachema, Czech Republic).
The activity of acid formation of bifidobacteria was determined by alkalimetric titration. To this end, 2 drops of phenolphthalein indicator were added to the daily cultures of bifidobacteria grown in Blaurok's medium in a volume of 5 ml and titrated with 0.1 M NaOH. The volume of sodium hydroxide that was used for titration was equimolar to the amount of the acid produced by microorganisms contained in 10 ml of the culture liquid. The result was expressed in Turner degrees:
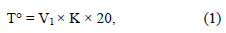
where V1 was the volume of a 0.1 M sodium hydroxide solution that was used to titrate 10 ml of the studied liquid, K was the correction factor determined when titrating a 0.1 M alkali solution with a 0.1 M succinic acid, and T° – the value that expressed the amount of
0.1 M sodium hydroxide used to titrate 100 ml of the studied sample.
The hydrophobicity of bifidobacteria was estimated according to Rosenberg et al. in the modification by L-Q Wang et al. To this end, the bifidobacteria were grown for 24 hours in a liquid Bifidum-medium (Obolensk) and then centrifuged at 8,000 g for 10 minutes. The bacterial mass was washed twice with a phosphate buffer and resuspended in the same solution. The optical density (A) of the suspension was determined at a wavelength of 600 nm. Then, 1 ml of dodecane was added to 3 ml of the bacterial suspension. The phases were mixed using Vortex for 2 minutes and left for 1 hour at 37°C to separate them. The optical density (A) of the aqueous phase was determined at 600 nm. The affinity to hydrocarbons was calculated as the percentage of hydrophobicity using the formula
![]()
where H is hydrophobicity, %; A0 and A are the optical density before and after processing the bacterial suspension by dodecane. The strains were considered highly hydrophobic at H = 60% and higher, mid- hydrophobic at H = 40–59%, and low-hydrophobic at Н ≤ 39%.
The adhesive properties of microorganisms were studied according to a method by V.I. Brilis (1986). To this end, slant meat-and-peptone agar cultures were grown for 24 hours. The suspension of microorganisms was prepared on the basis of a sterile isotonic sodium chloride solution at a concentration of 109 CFU/ml. The cellular substrate was the formalized human erythrocytes 0 (I) of the Rh (+) group, with a density of 100 million/ml. The erythrocytes and the suspension of microorganisms were combined in the equal volumes of 50 μl in test tubes and incubated at 37°C for 1 hour, regularly shaking the mixture. After that, a smear was prepared, dried, preserved in 96% alcohol for 15 minutes, and sent for Romanowsky-Giemsa staining. The adhesion was studied under a light microscope, the counting was carried out using 50 erythrocytes. The results of the experiment were estimated according to the microorganism adhesion index (MAI) that characterized the average number of microbial cells on one erythrocyte participating in the adhesive process. The microorganisms were considered non-adhesive at MAI ≤ 1.75; low-adhesive – from 1.76 to 2.5; medium- adhesive – from 2.51 to 4.0; and highly adhesive at MAI ≥ 4.0.
The lipoteichoic complex was extracted from the cell wall as follows. The bacterial culture grown in a liquid nutrient medium was washed, suspended in ice water and destroyed by ultrasonic treatment at 25 kHz. The suspension was centrifuged at an acceleration of 50,000 g for 15 minutes. The cell wall suspension was heated in water at 80°C for 3 minutes, then washed in water at 4°C. The non-covalently bound membrane protein was removed from the suspension by adding the equal volume of 5% (w/v) sodium dodecyl sulfate (SDS) and mixed for 4 hours at room temperature. To remove SDS, the cell walls were washed six times, replacing the volume of water with intermediate centrifugation at 50,000 g for 10 minutes at 20°C. The extraction was carried out using 10% (w/v) trichloroacetic acid with heating the sample at 90°C. After centrifugation at 50,000 g, the precipitation was carried out by adding excess dimethyl ketone in the cold. The carbohydrates were purified by ion exchange gel-permeation chromatography. To this end, the samples dissolved in 2 ml of water were applied to the column (30 cm × 1.5 cm) of DEAE cellulose pretreated with acetate pyridinium (pH 5.3), then equilibrated with water and eluted with a mixture of 50 ml of water and 50 ml of pyridinium acetate (pH 5.3). The fractions were collected in 2 ml by means of a Diafrak-002 fraction collector. The yield of the carbohydrate component in the fractions was controlled refractometrically.
The UV spectra were obtained using a SF-2000 instrument within the range of 190–340 nm with a resolution of 1 nm compared to a cuvette with a pure solvent.
Fourier transform IR spectroscopy was performed by means of a FSM-1202 instrument (Infraspek, Russia) using a MATIR sampler with a ZnSe crystal. The study was carried out in the transmission mode, within the range of 4000–465 cm-1, with a resolution of 4 cm-1, and a number of scans of 20. An empty cuvette was used as an example of comparison, the reference sample had been recorded immediately before each sample was analyzed.
In the gel-permeation chromatography (GPC) mode, Sephadex LH-20 (Pharmacia, Sweden) packed in a chromatographic column d = 6 mm was used as a chromatographic polysaccharide matrix (Pharmacia, Sweden). The volume of the adsorbent gel was 30 ± 1 ml, the elution rate was 0.1–0.15 ml/min, and the volume of the prepared fractions was 0.5 ml. The fractions were detected at a wavelength λ = 265 and 280 nm. Bidistilled water was used as an eluent.
The analysis was performed using the HPLC chromatograph "Tsvet Yauza-04" (with a UV and amperometric detector). The control of the instrument and the processing of the obtained results were carried out using the MultiChrome software, Version 3.1.1550 (ZAO Ampersend, Russia). The following chromatographic columns were used in this work: Gemini 5μm C18, 110A, 250 × 4.6 mm (Phenomenex, USA), Asahipak ODP-40 4E (4.6 × 250 mm). In terms of the trap column, the loop volume was 20 μl, the flow rate was 0.5 ml/min, and the pressure was 45.5 ± 1 bar. The mobile phase was made of acetonitrile for HPLC (Panreac); isopropyl alcohol for HPLC (Panreac); orthophosphoric acid (Panreac); and the bidistilled water prepared on the eve of the analysis. The components of MP in the volume ratio of 35:10:1:54 or 35:65 were mixed, and degassing was carried out using a vacuum aspirator.
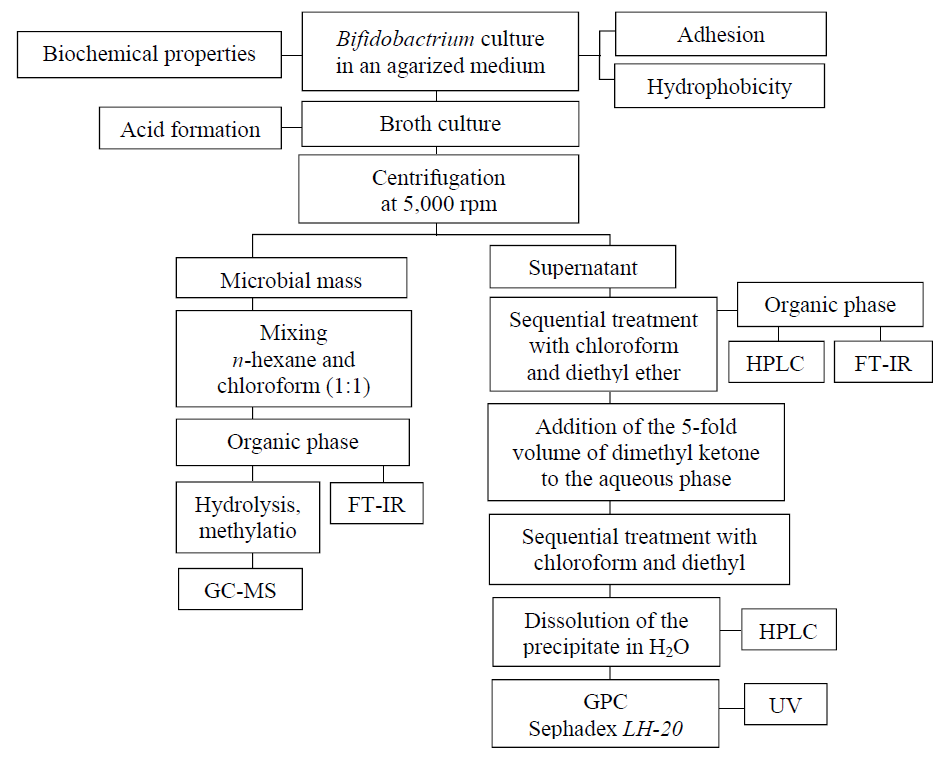
The fatty acid composition of bacterial lipids was determined by gas chromatography. To this end, the lipid fraction was isolated from a 0.9% broth culture of bifidobacteria preliminarily washed with a NaCl solution. The extraction was carried out using a chloroform and n-hexane mixture in a ratio of 1:1. The extract was methylated. A sample of 1 ml in volume was placed in a 1.5 ml vial, the solvent was blown away with nitrogen to dryness. 500 μl of a 3% of H2SO4 solution in MeOH was added to the dry residue. An internal standard element (10 μg of undecenoic acid) was added to the obtained solution. The sample was then heated at 90°C for an hour. After this, 700 μl of hexane was extracted. The volume of the hexane fraction was concentrated by stripping the solvent to a volume of 200 μl. The resulting sample containing fatty acids in the form of methyl ethers was used for analysis. The methylated samples were analyzed using an Agilent 7000B chromatomass spectrometer. The sample volume was 2 μl, splitless injection. Column: ZB-WAX, 30 m × 0.25 mm × 0.25 μm. Fig.1 presents the study design.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Some of bifidobacteria probiotic species from commercial preparations were used in the study. Various physicochemical characteristics that make it possible to determine the characteristic properties of probiotic species under in vitro conditions were compared. In general, there are several approaches that allow for the standardization of probiotic strains. The first approach is biological. It makes it possible to monitor the growth of a species as a whole when introducing a culture into the body, as well as to monitor the joint growth of a culture from a biological object or product/species in a selective growth medium. The count of colonies or turbidimetric measurements are used to determine the degree of inhibition of a pathogenic or conditionally pathogenic microflora using probiotic cultures. An alternative was the diffusion analyses used to estimate the inhibition of antagonistic strains [3]. On the other hand, at present, the physicochemical methods for standardization are being actively developed, which make it possible to supplement or obtain the detailed information on the properties of a probiotic strain and its metabolites.
The most important property of probitics is their ability to adhere to enterocytes. By attaching to the epithelium, microorganisms form a biofilm and thus provide competitive colonization resistance with pathogenic and opportunistic microorganisms. There are two groups of mechanisms of attachment of bacteria to cells or substrates: nonspecific and specific ones. Nonspecific adhesion is reversible and is related to the physicochemical features of bacterial cells – the total surface charge, hydrophobicity and the presence of ionic bonds with ionogenic groups. Nonspecific adhesion determines the primary interaction between a microorganism and the intestinal mucosa. According to modern concepts, hydrophobicity is considered as a property that allows bacteria to interact with organic molecules, form interassociative linksб and contact with the intestinal mucosa and immunocompetent cells.
In many respects, the hydrophobicity of a cell is determined by the lipid-like structures and, in particular, the fatty acids (FA) of the phospholipids of the cytoplasmic bacterial membrane. It is known that, regardless of the degree of hydrophobicity, saturated FA predominate in the cell wall of all the probiotic strains of bifidobacteria. In highly and low- hydrophobic strains, the proportion of saturated FA accounted for 64.2% and 71.3%, respectively, whereas, in the case of mid-hydrophobicity, the proportion of these acids was the highest and amounted to 91.1%. Table 1 presents the quantitative content of various FA in bifidobacteria. Approximately 90% of FA is used by microorganisms for the synthesis of phospholipids of the plasma membrane, 10% participate in the formation of lipoic acid, biotin, and in gram-negative bacteria (for the synthesis of lipopolysaccharides). Acyl carrier protein (ACP) plays a key role in the synthesis of fatty acids. Being a carrier that forms an acyl chain, it delivers precursor molecules from one enzyme to another for a condensation reaction. The enzymes of FA synthesis are localized in bacteria between the cytoplasm and the inner side of the plasma membrane. Their activity is determined by external factors, which determines the difference in the ways of FA synthesis. In this regard, the relative content of various fatty acids, the length of their chains, and saturation change. It has been established that in anaerobic bacteria, the precursor of unsaturated FA (C16:1, C18:1) is C10- ACP, and their formation is stimulated by a decrease in temperature and the oxygen content (the way of anaerobic synthesis).
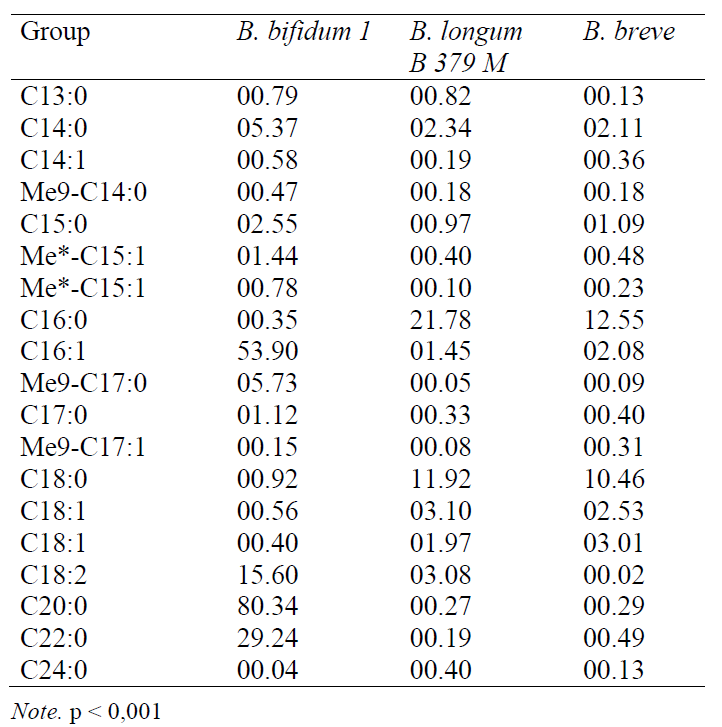
The aerobic way of synthesis of unsaturated FA from saturated precursors is well known and is characteristic of aerobic gram-positive bacteria. In the membrane of gram-positive bacteria, in addition to unbranched saturated FA, there are iso-, anteiso- and ώ-alicyclic FA that promote an increase in the plastic properties of the membrane. The synthesis of branched chain FA with a long carbon skeleton is characterized by the specificity of the enzymes that catalyze the condensation of molecules and the difference in the acyl carrier proteins that are seeds when the chain is elongated. Exogenous branched short chain carboxylic acids and branched 2-hydroxy acids are seeds. It should be emphasized that any changes in the microenvironment of bacteria will lead to differences in the ways of FA synthesis. This fact deserves special attention for study when choosing an optimal technological process when creating products with certain functional properties with a probiotic orientation. At the same time, it is important to determine the fatty acid composition of the membrane of the strains that differ from each other by hydrophobicity. The study of these factors will optimize the mechanisms of the positive effect of probiotics on a macroorganism.
In our study, the following saturated fatty acids were found in bifidobacteria using the GC-MS method (in order of an increase in the length of the carbon skeleton): n-tridecanoic (C13:0), n-tetradecane (myristic) (C14:0), n-pentadecane (C15:0), n-hexadecanoic (palmitic) (C16:0), n-heptadecane (C17:0) , n-octadecane (stearic) (C18:0), eicosanic (C20:0), behenic (C22:0), and lignoceric (C24:0) acids. Their quantitative content of which is presented in Table 1. Methylated acids or branched chain acids were also determined: 12-methyltetradecanoic (12CН3-С14:0), 13-methyl-tetradecanoic (13CН3-С14:0), isopentadecanoic (iso-C15:0), and isopalmitic (iso-C16:0). Branched or alicyclic FA, due to their structural features, perform an adaptive function in gram-positive bacteria giving fluidity and plasticity to the membrane [4]. Unsaturated FA, the highest proportion of which was found in bifidobacteria with high hydrophobic properties (35.8%), and, to a lesser extent, the strains with low hydrophobicity (28.7%) had similar properties. A low content of unsaturated FA was determined in the case of mid-hydrophobicity: their share did not exceed 8.9% in the whole FA pool. Among the monounsaturated FA, the bifidobacteria contained myristoleic (C14:1), pentadecenic (C15:1), palmitoleic (C16:1), n-heptadecene (C17:1), and oleic (C18:1) acids. There were also found diene FA: hexadecadiene (C16:2). In highly hydrophobic bifidobacteria, branched FA - isopentadecane (iso-C15:0) and methyl tetradecanoic (13CH3-C14:0) acids were identified. With the mid-hydrophobicity, a high content of n-hexadecane (iso-C16:0) and n-octadecane (C18:0) acids were established.It is known that the presence of unsaturated and branched FA in bacteria increases the fluidity of the membrane and promotes an increase in adhesion [14]. Indeed, the obtained data indicate the increased content and diversity of unsaturated FA with one or two double bonds in bifidobacteria with high hydrophobicity. In addition, in the case of high hydrophobicity, such branched fatty acids as isopentadecanoic (iso-C15:0) and 13-methyl-tetradecanoic (13CH3-C14:0) were found in bifidobacteria. In the strains of bacteria with the mid-hydrophobicity, only two unsaturated acids – oleic (C18:1) and linoleic (C18:2) – were present in relatively small amounts. In our opinion, the mid- hydrophobicity in the studied strains of bifidobacteria is apparently related to the presence of iso-palmitic (iso-C16:0) acid, the content of which was 20 times higher than that of the strains with high hydrophobicity. In the case of the low hydrophobicity of bacteria, the liquid-crystal state of the membrane was determined by unsaturated FA, the content of which was reduced in comparison with highly hydrophobic cultures.
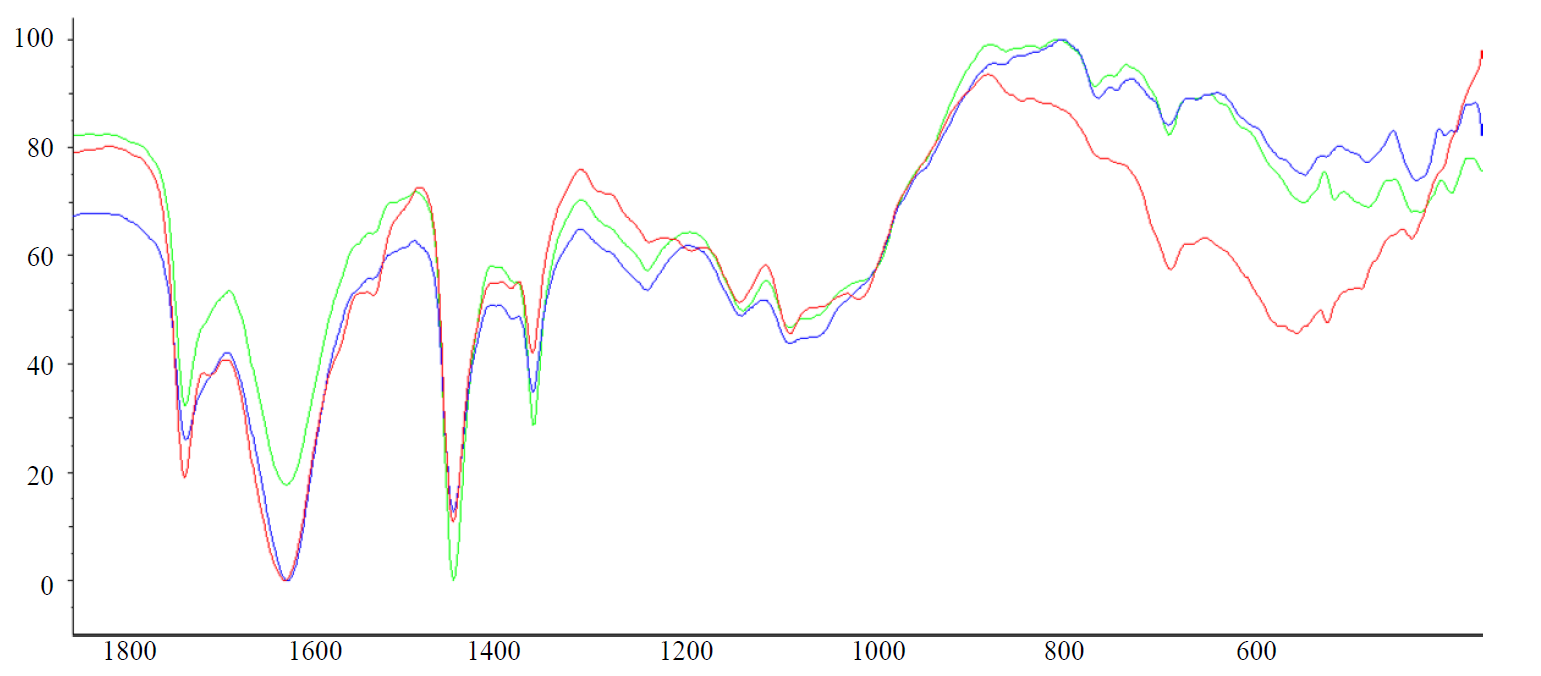
A large number of specific adhesion factors is a characteristic property for the representatives of the genus Bifidobacterium. These include stack-like structures, surface proteins, as well as lipoteichoic acids. They were found in a lot of gram-positive bacteria and consisted of 1,3-linked chains of polyglycerol phosphate and glycolipid fragments, but these polymers are so unique that they are used for the species identification of microorganisms. Specific adhesion is irreversible and takes place in microorganisms involving adhesin molecules (ligands), which have a high affinity for skin receptors and human mucosa. When studying the indicators of the specific adhesion of probiotic strains in vitro on erythrocytes, it was established that, regardless of the species, they had a mid ability for specific adhesion. Thus, MAI for B. bifidum 1 was 2.8, for B. breve – 3.1, and for B. longum В 379 М – 3.5. However, it is possible to judge specific adhesion indirectly using red blood cells as a model, since, despite the similarity of the glycoprotein receptors of erythrocytes and the intestinal epithelium, adhesion is tissue-specific. In this connection, the molecular characteristics of explicitly lipoteichoide-type adhesins in bifidobacteria were studied using FT-IR spectroscopy.
For the studied samples isolated from probiotic strains, the bands corresponding to the following structural fragments are determined. The presence of a 2952 cm-1 band is due to the stretching vibrations of C ̶ H and alkyl fragments. The asymmetric stretching vibrations (νasCH2) and symmetric (νsCH2) stretching vibrations determine the bands of 2,923 and 2,854 cm-1methylene groups. The presence of a 1738 cm-1 band characterized by an ether fragment is characteristic for all the samples under study. The strain B. bifidum 1 was the only that showed a weak intensity band of 1710 cm-1. The presence of this band, in our opinion, is a consequence of the structural features of adhesins in this type of bifidobacteria. These are the electronic effects of the nearest substituents, the compounds with multiple bonds, as well as the presence of inter- and intramolecular hydrogen bonds [15]. On the other hand, the presence of this band is explained by th possible effect of the methyl fragments located in the alkyl fragments, which is confirmed by the results of GC-MS [14]. Considering from these perspectives the shift of the absorption band of the carbonyl group, it is necessary to note the presence of unsaturated fragments in the structure of lipoteichoic acids which leads to the shift of absorption bands to the segment of the lowest frequencies. However, the effect of the O = P – OH fragment, which is part of the structure of fatty acids and which develops within the range of 1740–1600 cm-1, is the most probable [16]. Considering this band as a result of the effect of a phosphorus-containing fragment, one can find the confirmation of the presence of similar structures in other areas of the IR spectrum. Thus, for the sample of B. bifidum 1, the presence of the bands 1217, 1043 and 875 cm-1 is characteristic. On the basis of the given data of IR spectra and the established structural features, it should be recognized that the probiotic strain B. bifidum 1 is distinguished by a rich organophosphorus constituent. This structural feature positively influences the synthesis of highly ergic compounds and the metabolism of bacteria as a whole. Indeed, a wide spectrum of saccharolytic enzymes were determined in the study of the biochemical properties of this strain. B. bifidum 1 was able to ferment glucose, lactose, melibiose, raffinose, arabinose, xylose, galactose, fructose, and trehalose. The acid-forming capacity of the strain was 185 T°. Thus, the structural features of organophosphorus fragments, fixed under conditions of FT-IR spectroscopy, can be an informative criterion not only for the ability of bacteria for adhesion, but also for the metabolic activity of probiotic strains.
In turn, a relatively low variability of the spectral parameters found in the sample of B. breve should be noted (Fig. 2).
Among the positive impacts of probiotics, much attention is paid to exometabolites, which create favorable conditions for the growth and reproduction of one's own human microflora and modulate its metabolic processes. Therefore, it is expedient to develop criteria for the estimation and standardization of exometabolites of probiotic microorganisms.
To study the culture liquid of bifidobacteria, liquid column chromatography was used with Sephadex LH-20 dextran sorbent. The molecular weight fractionation range for LH-20 was 100 for the lower limit and from 2,000 to 10,000, according to some data, from 4,000 to 6,000, for the upper limit [17]. In most cases, the peptides with more than five or seven amino acid residues are soluble only in polar solvents, so it is optimal to perform their separation and preparative accumulation by gel filtration using soft dextran sorbents such as LH-20, stable in organic solvents [18, 19]. Moreover, it should be noted that the pattern of separation using Sephadex LH-20 is determined not only by gel filtration but also by adsorption and distribution chromatography, and the effective fractionation interval depends both on the eluent used and the conformation of the fractionated substances [20]. According to the analytical data obtained in environments of ion- exclusion chromatography and GC-MS, the studied exometabolites did not contain any nucleic and organic acids. There were no pentoses in any of the hydrolysates of exometabolites.
The product is free of lipids according to the results of gas chromatography after extraction with chloroform and diethyl ether. The results obtained by high-performance liquid chromatography of HPLC using an Asahipak ODP-40 4E column showed that the product had a glycoprotein (high molecular weight) nature. The exopolymer produced by B. bifidum 1, B. longum 379 M, and B. breve strains consisted mainly of glucides (measured using the phenolic- sulfuric acid method) and proteins (measured using Lowry's method) in a ratio of 5:1.
The glucid part consisted of glucose and galactose according to the analytical data obtained by ion- exclusion, thin-layer and gas chromatography. Glucose and galactose were present in a ratio of 3:2. According to the results of the gas chromatography of the monomers hydrolyzed by methanolysis and then acetylated with trifluoroacetylation, the polysaccharide part had a variety of bonds, such as 1–2, 1–3, 1–4, and 1–6.
The molecular-mass distribution (MMD) of the exometabolites of the studied strains in the chromatography mode using Sephadex LH-20 was indicated by the following features. The spectrum of the metabolites produced by bifidobacteria was quite wide. In the case of GPC, in the mode of water elution in a volume of 15–17, the yield of the component c in the UV segment λ max = 243 nm was noted in the sample of B. breve (2) (Fig. 4c). In our opinion, this spectral characteristic is determined by the presence of steroid-like structures. The yield of the component with a lower MMD of this sample was in a volume of the fraction 23–25 and has a UV spectrum with a maximum of 265 nm (Fig. 4d). As is known, such spectral characteristics are typical for protein-peptide structures. It is noteworthy that this component of B. bifidum 1 using Sephadex LH-20 sorbent elutes in the fraction zone 21–27 as a single chromatographic peak. The spectrum in the UV segment had a maximum of 265 nm (Figs. 4a and b). The sample of exometabolites isolated from the culture medium B. longum B 379 M had the highest heterogeneity of MMD components. The chromatographic profile of this sample was represented by quite well separated peak fractions (3) (Fig. 3). The UV spectrum of the components eluted in the zone of the fraction 27–31 characterized the presence of a maximum of absorption of 264 nm, whereas this value was 272 nm for fractions 37–45 (Fig. 4e). These features should be considered as the distinctive conformational features of protein- peptide components in the presence of lipoprotein formations.
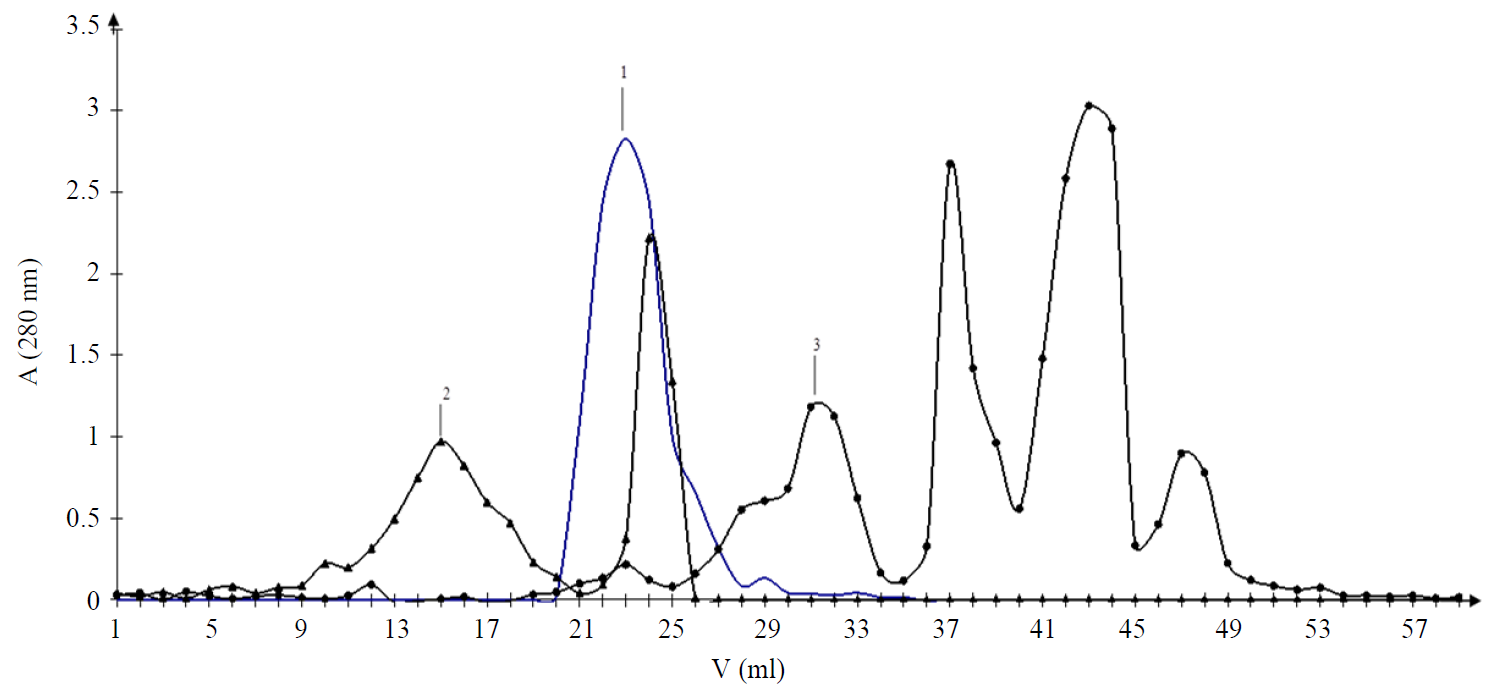
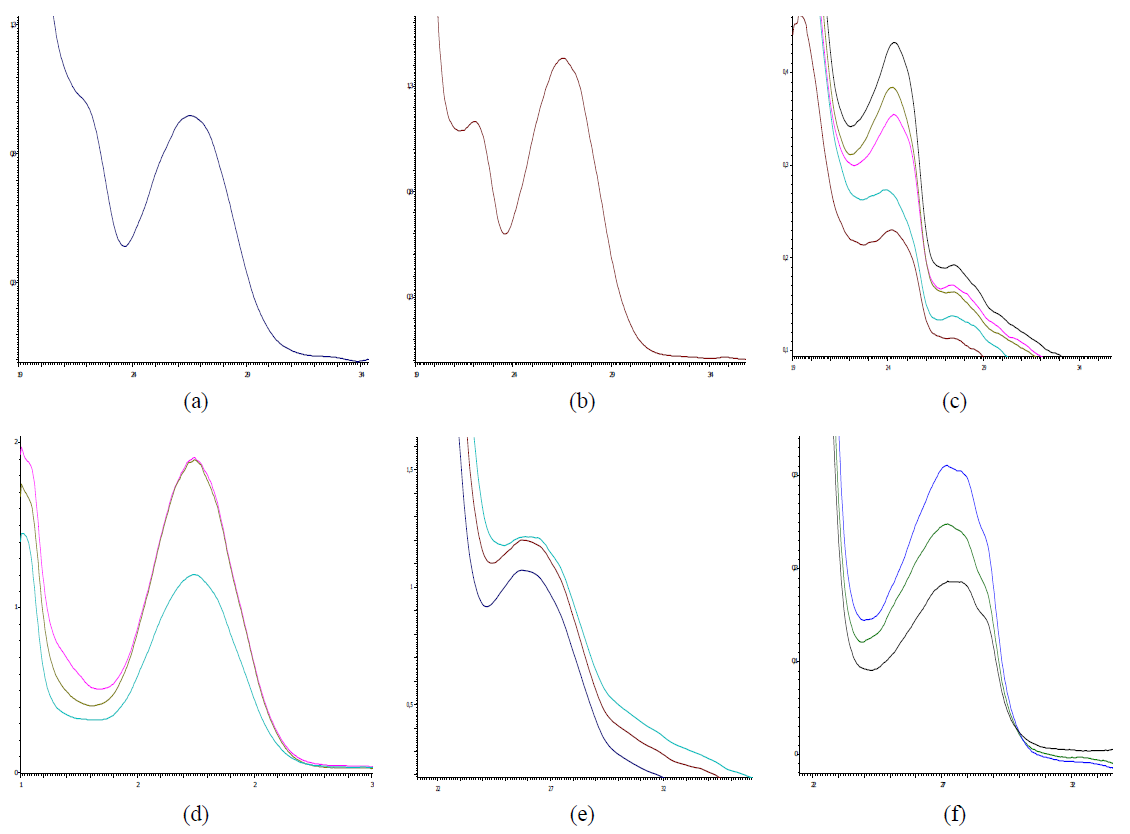
Thus, in the mode of GPC, using Sephadex LH-20, a fraction with a low degree of MMD and quite homogeneous UV spectra was released from the sample of B. bifidum 1. The feature of B. breve was the variability of the spectral indices. The effect of MMD under conditions of GPC, in turn, is the significant indicator of the enzymatic activity of the studied strains of microorganisms. As it was established, the strain B. bifidum 1 had a low enzymatic activity, in turn, the strains B. longum 379 M and B. breve had a wide set of saccharolytic enzymes and were capable of fermenting various carbohydrates [10]. In the development of probiotic products of functional nutrition, strains with a low enzymatic activity are often used in multicomponent bacterial consortia to create favorable conditions for their joint cultivation and to preserve the viability of all participants in a multicomponent community. Indeed, the B. bifidum 1 strain was isolated from a two-component probiotic consortium, so GPC allowed estimating not only the enzymatic activity of the strain, but also making an assumption about the number of strains in a functional product. The results of MMD obtained in our study allow us to say that the B. longum B 379 M strain has a relatively high enzymatic activity. Under conditions of GPC, using Sephadex LH-20, the analyte of exometabolites could be fractionated into four chromatographic zones, whereas the sample of B. longum B 379 M is characterized by the presence of fractions with the distinctive parameters of MMD and/or conformation.
The use of the GPC version in combination with the study of the spectral parameters of fractions in the UV segment also reveals the distinctive physicochemical characteristics of exometabolites of bifidobacteria strains. The exopolysaccharides produced by microorganisms are most often the basis for feeding other members of the normal intestinal microflora (Bifidobacterium, Escherichia coli), which stimulates its growth, development, and metabolic activity, i.e., the exometabolites of a polysaccharide nature have prebiotic properties. The produced exopolymer matrix also promotes the formation of a full biofilm on the intestinal mucosa. Like a "glove", it covers the intestinal mucosa and mechanically protects the intestinal biotope from being populated by pathogenic microorganisms. In this case, the minor amounts of the produced protein level out the possibility for microorganisms to multiply in the biotope with a proteolytic activity and minimize the development of putrefactive dyspepsia. This feature is especially relevant in the development of specialized probiotic products for high-risk groups [21]. The use of sorption- chromatographic approaches in the analysis and study of biofilm formation in probiotic strains appears to be promising.
Thus, in our opinion, it is expedient to select the strains of microorganisms that are promising for the biotechnological process and also to estimate the functional characteristics of the applied probiotic cultures using a set of classical bacteriological and physico-chemical methods. The most informative chromatography methods for studying the factors of specific and nonspecific adhesion and exometabolites are GC-MS and GPC, as well as FT-IR spectroscopy.
To estimate the non-specific adhesion of probiotic strains, it is expedient to use the data on the fatty acid composition of the cell walls of bifidobacteria. The criterion for the high ability of strains for reverse adhesion is the predominance of the carbon skeleton and unsaturated fatty acids in the cell wall of fatty acids with a branched structure.
The estimation of structural features of lipoteichoic acids using IR spectroscopy allows us to estimate not only the ability of probiotic cultures for specific adhesion, but also their functional potential. The indicator of high biological activity of strains is the presence of a large number of phosphoric acid fragments in lipoteichoic acids.
The most informative estimation of the composition of the exometabolites produced by probiotic bifidobacteria is provided by a complex of such chromatographic methods as GC-MS and GPC, as well as FT-IR spectroscopy. The predominance of bifidobacteria of glucides in the composition of exometabolites is an indicator of the high saccharolytic activity of probiotic strains.
КОНФЛИКТ ИНТЕРЕСОВ
The authors declare no conflict of interest.
ФИНАНСИРОВАНИЕ
The study was supported by the Kemerovo State Medical University, the Russian Federation Ministry of Health.
СПИСОК ЛИТЕРАТУРЫ
- Vinderola G., Gueimondeb M., Gomez-Gallego C., Delfederico L., and Salminen S. Correlation between in vitro and in vivo assays in selection of probiotics from traditional species of bacteria. Trends in Food Science and Technology, 2017, vol. 68, pp. 83–90. DOI: https://doi.org/10.1016/j.tifs.2017.08.005.
- Dodooa C.C. Wanga J., Basitab A.W., Stapletona P., and Gaisforda S. Targeted delivery of probiotics to enhance gastrointestinal stability and intestinal colonization. International Journal of Pharmaceutics, 2017, vol. 530, no. 1–2, pp. 224–229. DOI: https://doi.org/10.1016/j.ijpharm.2017.07.068.
- Fredua-Agyeman M., Stapleton P., Basit A.W., and Gaisford S. Microcalorimetric evaluation of a multi-strain probiotic: Interspecies inhibition between probiotic strains. Journal of Functional Foods, 2017, vol. 36, pp. 357–361. DOI: https://doi.org/10.1016/j.jff.2017.07.018.
- Zakharova Yu.V., Otdushkina L.Yu., Levanova L.A., and Sukhikh A.S. Mechanisms of bifidobacteria resistance to lipolytic enzymes of Staphylococcus aureus. Fundamental and clinical medicine, 2017, vol. 2, no. 1, pp. 6–13. (In Russ.).
- Tripathi M.K. and Giri S.K. Probiotic functional foods: Survival of probiotics during processing and storage. Journal of Functional Foods, 2014, vol. 9, no. 1, pp. 225–241. DOI: https://doi.org/10.1016/j.jff.2014.04.030.
- Hossain Md.I., Sadekuzzaman M., and Ha S.-D. Probiotics as potential alternative biocontrol agents in the agriculture and food industries: A review. Food Research International, 2017, vol. 100, part 1, pp. 63–73. DOI: https://doi.org/10.1016/j.foodres.2017.07.077.
- Daliri E.B.-M. and Lee B.H. New perspectives on probiotics in health and disease. Food Science and Human Wellness, 2015, vol. 4, no. 2, pp. 56–65. DOI: https://doi.org/10.1016/j.fshw.2015.06.002.
- Dwivedi M., Kumar P., Laddha N.C., and Kemp E.H. Induction of regulatory T cells: A role for probiotics and prebiotics to suppress autoimmunity. Autoimmunity Reviews, 2016, vol. 15, no. 4, pp. 379–392. DOI: https://doi.org/10.1016/j.autrev.2016.01.002.
- Biancone L., Vavassori P., Monteleone I., et al. Cytoskeletal proteins and resident flora. Digestive and Liver Disease, 2002, vol. 34, no. 2, pp. 34–36. DOI: https://doi.org/10.1016/S1590-8658(02)80161-X.
- Kailasapathy K. and Chin J. Survival and therapeutic potential of probiotic organisms with reference to Lactobacillus acidophilus and Bifidobacterium spp. Immunology and Cell Biology, 2000, vol. 78, no. 1, pp. 80–88. DOI: https://doi.org/10.1046/j.1440-1711.2000.00886.x.
- Mc Kean J., Naug H., Nikbakht E., Amiet B., and Colson N. Probiotics and Subclinical Psychological Symptoms in Healthy Participants: A Systematic Review and Meta-Analysis. The Journal of alternative and complementary medicine, 2017, vol. 23, no. 4, pp. 249–258. DOI: https://doi.org/10.1089/acm.2016.0023.
- MU 2.3.2.2789-10. Metodicheskie ukazaniya po sanitarno-epidemiologicheskoy otsenke bezopasnosti i funktsional'nogo potentsiala probioticheskikh mikroorganizmov, ispol'zuemykh dlya proizvodstva pishchevykh produktov [Methodical instructions 2.3.2.2789-10 Methodical instructions for the sanitary-epidemiological assessment of the safety and functional potential of probiotic microorganisms used for the production of food products]. Moscow, 2011, 105 p.
- Gill H.S. and Guarner F. Probiotics and human health: a clinical perspective. Postgraduate Medical Journal, 2004, vol. 80, no. 947, pp. 516–526. DOI: https://doi.org/10.1136/pgmj.2003.008664.
- Zakharova Yu.V. and Sukhikh A.S. Chromatographic analyses of membrane fatty acid Bifidobacterium with different hydrophobicity. Sorption and chromatographic processes, 2015, vol. 15, no. 6, pp. 776–783. (In Russ.).
- Silverstein R.M., Webster F.X., and Kiemle D.J. Spectrometric identification of organic compounds. Hoboken, NJ: John Wiley & Sons, 2005. 551 p.
- Stuart B.H. Infrared spectroscopy: fundamentals and applications. Hoboken, NJ: John Wiley & Sons, 2004. 242 p.
- Sukhikh A.S. and Kuznetsov P.V. Application of a sorbent of universal appointment Sephadex LH-20 in modern medical and biologic researches. Medicine in Kuzbass, 2009, vol. 8, no. 4, pp. 3–12. (In Russ.).
- Borisenko V., Lougheed T., Hesse J. Simultaneous optical and electrical recording of single gramicidin channels. Journal Biophysical, 2003, vol. 84, pp. 612–622.
- Nefedov P.P., Zhmakina T.P. and Belenkiy B.G. Vysokoeffektivnaya eksklyuzionnaya zhidkostnaya khromatografiya zashchishchennykh peptidov na myagkikh i poluzhestkikh gelyakh [Highly efficient exclusive liquid chromatography of protected peptides on soft and semi-rigid gels]. Russian Journal of Bioorganic Chemistry, 1983, vol. 9, no. 5, pp. 616–627. (In Russ.).
- Sussman J.L., Barzilay I., Keren-Zur M., and Lapidot Y. Correlation of the differences in conformation between 2′–5′ and 3′–5′ dinucleoside monophosphates with their behaviour on a Sephadex LH-20 column. BBA Section Nucleic Acids and Protein Synthesis, 1973, vol. 308, no. 2, pp. 189–197. DOI: https://doi.org/10.1016/0005-2787(73)90148-2.
- Sanders M.E., Merenstein D.J., Ouwehand A.C., et al. Probiotic use in at-risk populations. Journal of the American Pharmacists Association, 2016 vol. 56, no. 6, pp. 680–686 DOI: https://doi.org/10.1016/j.japh.2016.07.001.
